Entsteht das Denken im Gehirn? Wie gut lassen sich Nahtoderfahrungen und andere spirituelle Erfahrungen im Rahmen materialistischer, quantenphysikalischer und spiritueller Deutungssysteme verstehen?
Wie man dem Rest meiner Internetseite entnehmen kann, zähle ich zu den Leuten, die der Ansicht sind, daß das Bewußtsein nicht im Gehirn entsteht. Nichts desto trotz, wurde in der Wissenschaft vor allem zwischen 1900 und 2000 vorrangig mit der Hypothese gearbeitet, daß Materie das Primäre wäre und das das Gehirn eine Art Denkmaschine ist. Es wurden auch sehr viele Belege zusammengetragen, die sehr gut in dieses Modell paßten.
Schon fast so lange wie diese Geräte existieren, wurden Radio und Fernsehen als Analogie herangezogen, um zu erklären, warum man diese Beobachtungen auch anders deuten kann. Eine prägnante Formulierung dieses Arguments bringt Stanislav Grof:
Der Fernseher als Analogie um das Gehirn als Informationsempfänger zu verstehen
Im Falle von begrenzten Gehirntumoren kann der funktionelle Schaden - Sprachverlust, Verlust der motorischen Kontrolle und so fort - dazu dienen, genau zu diagnostizieren, welche Stelle den Gehirns betroffen ist.
(...)
Ein guter Fernsehmechaniker kann uns anhand der fehlerhaften Bild- und Tonwiedergabe ganz genau sagen, was mit dem Apparat los ist und welche Teile ersetzt werden müssen, damit er wieder richtig läuft. Aber niemand würde deshalb behaupten wollen, der Fernseher sei selbst für die Programme verantwortlich, die wir sehen, wenn wir ihn anstellen.
1. S.17
Natürlich gibt es noch die dritte Möglichkeit, daß das Gehirn Bewußtsein teilweise empfängt, teilweise aber auch produziert. Man muß also mit drei Analogien arbeiten.
Der Fernseher als reiner Informationsempfänger
Der Computer ohne Internetanschluß als Datenverarbeitungsgerät mit Datenspeichern
Das Smart-Phone, das sowohl Informationsempfänger ist, als auch Datenverarbeitung und Datenspeicher besitzt
Neben den Eigenschaften, die ich für diese drei Geräte oben aufgezählt habe, hat der menschliche Körper natürlich auch das Gegenstück einer Videokamera, mit dem man Farbfilme mit Ton aufnahmen kann und die Funktionen, mit denen ein Roboter ausgestattet ist, der in der äußeren Welt auch handeln kann.
Im Folgenden geht es darum, all diese Beobachtungen zu sammeln, zu prüfen, wie diese Beobachtungen im "Gehirn erzeugt denken"- und "Gehirn empfängt denken"-Modell verstehen lassen und welche der beiden Deutungen die beobachteten Fakten besser erklärt.
2.1 Grundgedanken zur Wahrnehmung, Gehirn und Informationsquelle
Eine Sinneswahrnehmung läuft grob in folgenden Schritten ab:
Wahrnehmung im Sinnesorgan
Nervenleitung
Verarbeitung im Gehirn
subjektiver Seheindruck
Prinzipell ist jeder dieser Schritte schon sowohl beim Mensch als auch bei unterschiedlich komplexen Tieren in verschiedenster Weise untersucht worden. Man weiß daher recht detailliert, wie das Gehirn bei der Sinneswahrnehmung arbeitet.
Es ist prinziepiell für diverse Sinne bei vielen verschiedenen Lebewesen nachgewiesen, wie der wahrzunehmende Reiz zu der Sinneszelle gelangt, wie die Sinneszelle chemisch funktioniert, um den Reiz wahrzunehmen, wie die Reizung der Sinneszelle an den Nerv weitergegeben wird, wie der Nerv funktioniert, um die Information zum Gehirn weiterzuleiten, an welchen Stellen die Wahrnehmung von einer Nervenzelle auf die andere übertragen wird, wo im Gehirn die Information ausgewertet wird und es sind auch einige Details darüber bekannt, wie das Wahrzunehmende ausgewertet wird. Wenn man sich einen groben Überblick verschaffen will, wie der Stand der Wissenschaft dazu ist, macht es Sinn ein aktuelles Lehrbuch zur Neurophysiologie, das ist sie Wissenschaft dazu, wie das Nervensystem funktioniert, zu lesen. Meines war beispielsweise im Studium "Neurowissenschaft"10.. Die komplexeren Auswertungsvorgänge komplizierterer Gehirne sind aber nicht vollständig verstanden.
Daneben gibt es noch diverse Möglichkeiten, bei denen subjektiv etwas wahrgenommen wird, das an eine Sinneswahrnehmung erinnert, das objektiv nicht mit dem zugehörigen Sinnesorgan registriert wird.
Bei verschiedenen Halluzinationen kann es passieren, daß wir Dinge wahrnehmen, die im Äußeren Raum nicht vorhanden sind.
Bei Illusionen hat das Wahrgenommene zwar ein Gegenstück in der äußeren Welt, es wird aber in irgendeiner Form falsch oder verzerrt wahrgenommen oder interpretiert
Wahrnehmungen, die durch techniche Geräte ins Gehirn eingespielt werden, z.B. Cochlea implantat
Feinstoffliche Wahrnehmungen (z.B. Telepathie, Wahrnehmungen durch eine Seele, die sich außerhalb des Körpers befindet, sehen von Geistern)
Während Halluzinationen, Illusionen, Fantasievorstellungen, Träume, Synästhesie und Wahrnehmungen die durch techniche Geräte ins Gehirn eingespielt werden prinzipiell im Rahmen eines Weltbildes erklärbar sind, das annimmt, daß das Gehirn das Bewußtsein erzeugen würde, stellen überprüfbar richtige feinstoffliche Wahrnehmungen dieses Weltbild je nach Art der Wahrnehmung vor unterschiedliche Probleme.
Fantasievorstellungen, Träume, Synästhesie und technisch eingespielte Wahrnehmungen zeigen uns, daß der Gehirnbereich, der beispielsweise zum sehen gehört, auch Informationen sinnvoll verarbeitet, die nicht vom Auge geliefert werden und daß wir diese Informationen zwischen sehr vage und fast so, als hätten wir es mit dem Auge gesehen, wahrnehmen können. Dies werde ich im Folgenden zum Vergleich verwenden, um genauer einzuordnen, wie feinstoffliche Wahrnehmungen einzuordnen sind.
Im Einzelnen versuche ich folgende Fragen zu klären
Gibt es Wahrnehmungen nachweisbar zutreffender Tatsachen, die sich nicht erklären lassen, indem man annimmt, das Wissen wäre durch die normalen Sinne oder sonstige materielle Hilfsmittel vermittelt oder durch logisches Denken erschlossen worden.
Wenn diese Frage mit Ja beantwortet wird ergeben sich daraus folgende Fragen
Wie unterscheidet sich feinstoffliche und irdische Wahrnehmung?
An welcher Stelle des Wahrnehmungssystems werden feinstoffliche Wahrnehmungen ins Gehirn eingespielt?
2.2 Das Sehen als Beispiel für eine Sinneswahrnehmung des Menschen
2.2.1 Das Auge als abstimmbares Linsensystem
Um beurteilen zu können, wie groß die Stärken und was die Schwächen einer Modellvorstellung sind, die annimmt, Bewußtsein und Denken würde durch das Gehirn produziert, braucht man eine grob richtige Vorstellung, wie viel wir über die Funktionsweise des Gehirns wissen. Während es nicht machbar ist, das im Rahmen dieses Textes für alle bekannten Gehirnfunktionen zu erledigen, will ich doch für einen Sinn einmal sehr ins Detail gehen, da wohl nicht einmal diejenigen, die Fachleute für einen Teilbereich dieses Wissens sind, einen vollständigen Überblick über alles haben, was es da zu wissen gäbe. Ich weiß natürlich auch nicht alles, aber ich habe schon in vielen Bereichen verschiedenster Wissenschaften sozusagen Probebohrungen unternommen, die mir zeigen, was man typischerweise findet, wenn in diesem Bereich zu bohren beginnt.
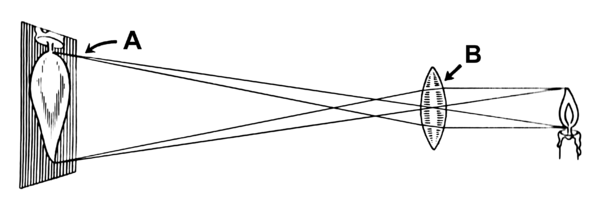
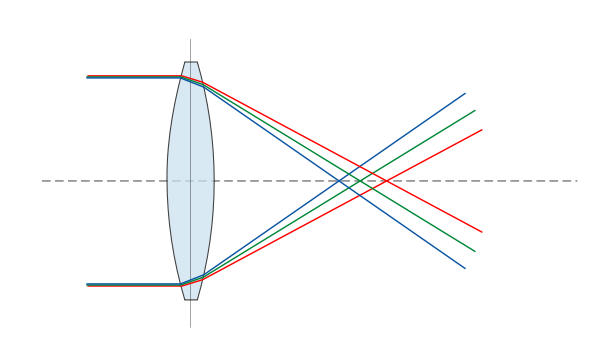
Abbildung einer Kerzenflamme (Objekt) mit einer Linse (B) auf eine Leinwand (A), als vergößertes umgekehrtes Bild (A) der Flamme
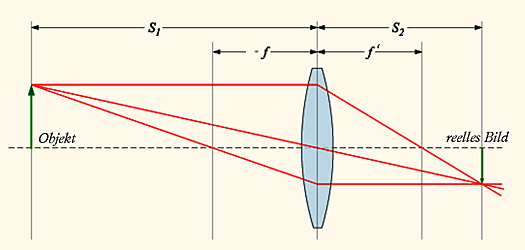
Mit einer Sammellinse, kann man ein Bild eines Gegenstandes auf einer Leinwand oder anderen Fläche erzeugen. Wo das Bild entsteht, hängt davon ab, wo sich der Gegenstand befindet und wie groß die Brennweite der Linse ist.
Die Brennweite f gibt an, wie weit hinter der Linse der Punkt liegt, an dem sich alle Strahlen treffen, wenn man ein paralleles Strahlenbündel auf die Linse treffen läßt. Ein fast paralleles Strahlenbündel ist das Licht der Sonne, wenn es auf der Erde auftrifft, so daß man dieses verwenden kann, um den Brennpunkt einer Linse zu bestimmen.
In normales Situationen hat man allerdings zunächst kein paralleles Strahlenbündem, sondern das Licht breitet sich von dem Gegenstand von dem es ausgeht in alles Richtungen aus. Wenn sie dann auf die Linse treffen geht der Strahl, der durch die Mitte der Linse geht geradeaus weiter, diejenigen die anfang parallel zur Achse des Systems liefen, gehen nachher durch den Brennpunkt und die, die am Anfang durch den Brennpunkt gelaufen sind,
Um zu berechnen, wie die verschiedenen Größen miteinander zusammenhängen
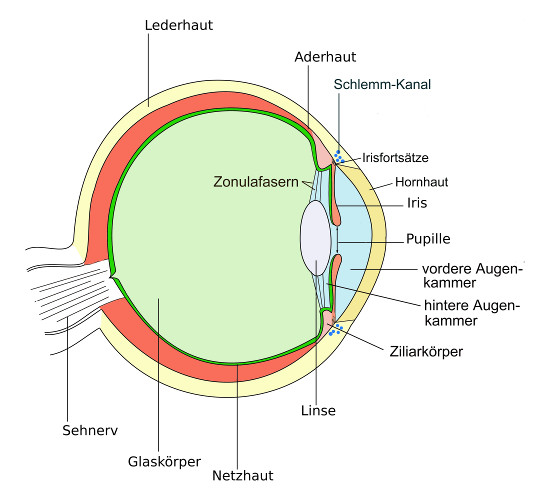
Texte dazu, wie das sehen funktioniert, beginnen gewöhnlich mit einer Darstellung wie der Folgenden, in der das Auge als abstimmbares Linsensystem verstanden wird, das den abzubildenden Gegenstand scharf auf der Retina abbildet, ähnlich wie ein Photoapparat so eingestellt wird, daß das Bild scharf auf dem Film oder den digitalen Fotosensoren abgebildet wird, die als Leinwand dienen.
Während beim Fotoapparat gewöhnlich der Abstand zwischen Linse und Film verstellt wird, wird im Auge die Linse nach der üblichen Betrachtungsweise mit der Muskelkraft des Ziliarmuskels im Ziliarkörper verformt. Es ändert sich also nicht der Abstand von Objekt und Bildes sondern die Brennweite der Linse.
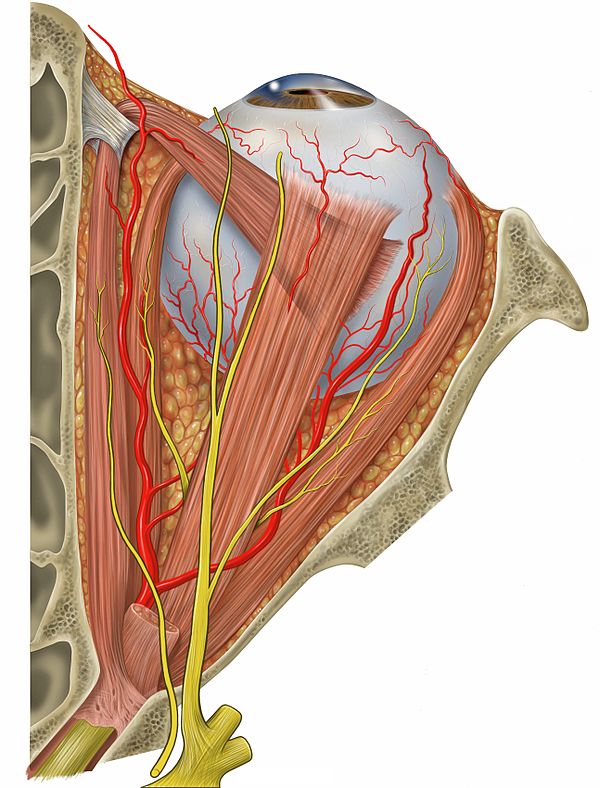
Blick auf den Augapfel mit den umgebenden Muskeln von oben
Am Augapfel setzen diverse Muskeln an, die dazu dienen, ihn so zu bewegen, daß wir genau das ansehen, was wir sehen wollen. William Horatio Bates vertrat die Ansicht, daß diese Augenmuskeln darüber hinaus auch den Augapfel so verformen, daß dadurch das Bild scharf gestellt wird12. S.30ff, 13. S.19f. Aus meiner Sicht sind Muskeln, die so am spürbar elastischen Augapfel ansetzen, wie diese Muskeln das tun, prinzipiell zusätzlich zu den üblicherweise genannten Funktionen in der Lage den Augapfel weiter in die Augenhöhle hineinzuziehen und ihn zu verformen. Die Behauptung, das wäre nicht der Fall, kommt mir daher nicht glaubhaft vor. Ob ein gesundes Arbeiten des Auges die Akkodomation hauptsächlich über den Ziliarmuskel bewerkstelligt oder hauptsächlich mit den Muskeln, die das Auge auch drehen und ausrichten und ob eine Verformung des Augapfels daher eher eine Fehlfunktion oder aber eine normale Funktion dieser Muskeln ist, läßt sich nach dieser Darstellung nicht sicher entscheiden.
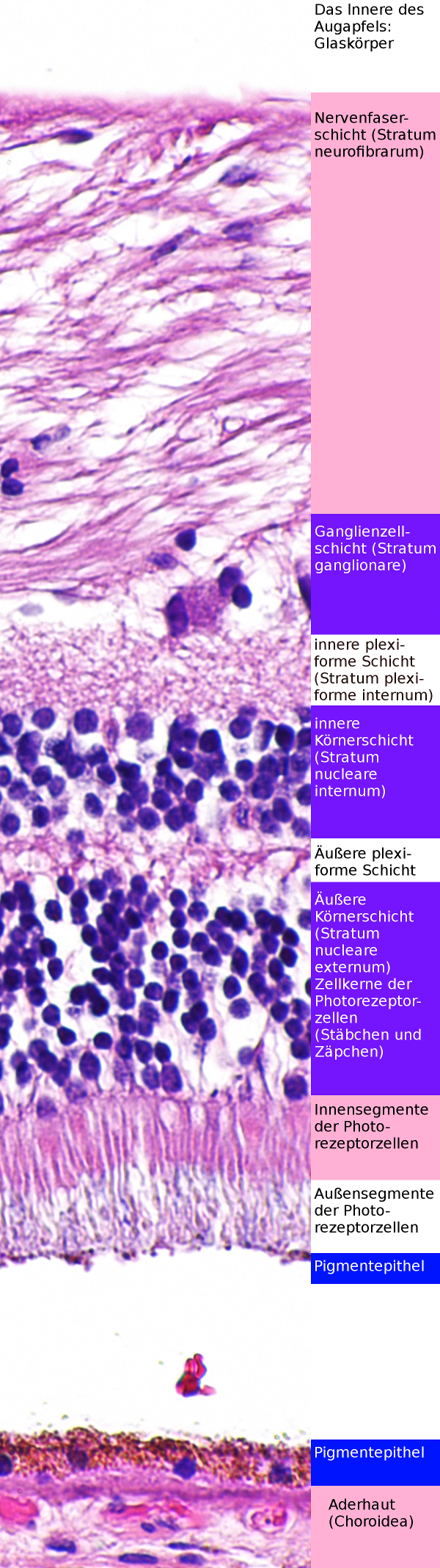
Netzhaut (Retina) nahe der Eintrittstelles des Sehnervs.
Die Nervenfaserschicht ist an dieser Stelle ausgesprochen dick. Der Riß im Pigmentepithel gehört da nicht hin - das Präparat ist an dieser Stelle zerrissen. H&E-Färbung
VA83.3.3
Färbungen im Durchlichtmikroskop
Oben ist die Netzhaut (Retina) in einem histologischem Schnitt (Histologie = mikroskopische Anatomie) zu sehen, unten ist dasselbe schematisch dargestellt. Wie man sehen kann, muß das Licht zuerst durch die Nervenfaserschicht, die Ganglienzellschicht, die innere und die äußere plexiforme und Körnerschicht hindurch, bevor es an die Außensegmente der Photorezeporzellen gelangt, in denen sich die Sehfarbstoffe befinden, die gebraucht werden, um das Licht wahrzunehmen.
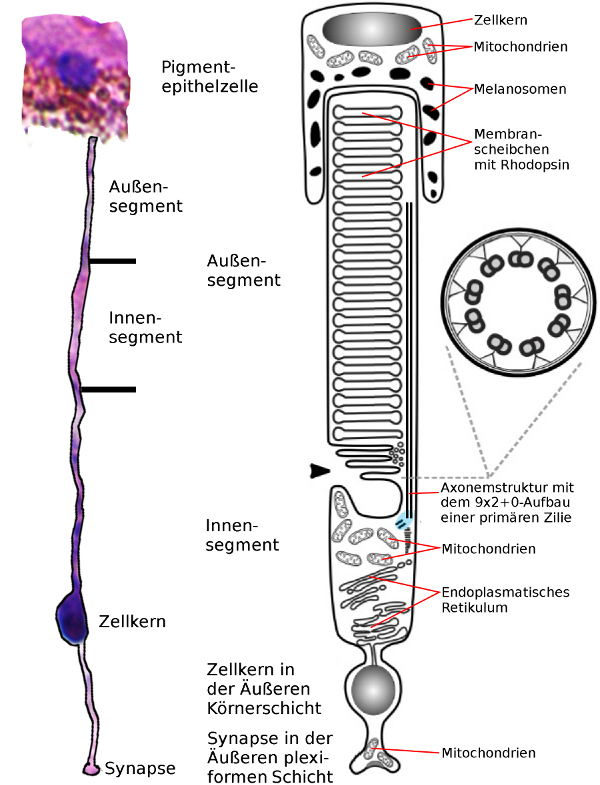
Das Stäbchen ist eine umgebaute Nervenzelle, deren primäre unbewegliche Zilie in das Äußere Segment umgewandelt wurde.
Daß äußere Segment des Stäbchens steckt mit der äußeren Spitze in einer Pigementzelle des Pigmentepithels, die verbrauchte Teile der Membranstapel phagozytiert. Das Äußere Segment enthält in seiner ganzen Länge Stapel aus dicht gepackten Membranscheibchen, in denen sich der Sehpurpur, das Rhodopsin befindet, ein Protein, das als Rezeptor zur Lichtwahrnehmung dient.
Zwischen dem inneren und den Äußeren Segment findet sich ein kurzes dünnes Stück, das als Skellett eine Struktur besitzt, die aus neun im Kreis angeordneten Paaren an Mikrotubuli besteht, als Axonemstruktur bezeichnet wird und typisch für Zilien ist. Diese Axonemstruktur reicht weit in das äußere Segment hinein und bildet dessen Rückrat, durchquert das dünne Verbindungsstück und endet im Basalkörper der Zilie, die die Grenze zum inneren Segment darstellt.
Das innere Segment des Stäbchens stellt den größten Teil des Zellkörpers der Photorezeptorzelle dar und enthält unter anderem Mitochondrien - die Kraftwerke der Zelle und das Endoplasmatisches Retikulum, an dem die Ribosomen sitzen, die die Proteine der Zelle produzieren - beispielsweise auch den Sehpurpur.
Der Zellkern befindet sich nicht im inneren Segment sondern er hängt an einem langen dünnen Stil und befindet sich irgendwo in der Äußeren Körnerschicht. Ein weiterer Stil verbindet den Zellkern mit der Synapse in der äußeren Plexiformen Schicht. Da die Zellkerne viel dicker sind als das Innensegment der Photorezeptorzellen müssen sie in mehreren Schichten angeordnet werden, damit alle Platz finden. Daher sieht die Körnerschicht aus, als waren da lauter unregelmäßig verteilte Körner, von denen jedes auf einem langen Faden aufgefädelt ist, der von dem Innensegment bis zur Äußeren plexiformen Schicht reicht.
Das Stäbchen ist hier genau umgekehrt dargestellt, wie im histologischen Schnitt oben. Das Innere des Auges ist also unten, wo die Synapse ist, der äußere Rand des Auges ist oben bei der Pigmentepithelzelle. Die Fadenförmige Verbindung zwischen Innensegment und Zellkern sowie zwischen Zellkern und Synapse sind beide sehr stark verkürzt dargestellt. Der Zellkern ist im Verhältis wesentlich größer als dargestellt.
Ein einzelnes Stäbchen habe ich aus der obigen Darstellung ausgeschnitten und daneben gelegt, um deutlich zu machen, wie die Größenverhältnisse wirklich sind. - wobei die Dicke wegen dem verschwommenen Originalbild nur sehr ungefähr hinhaut, während die Grenzen von Innen- und Außensegment richtig angegeben sind.
2.2.3.1.2 Das äußere Segment ist eine primäre Zilie
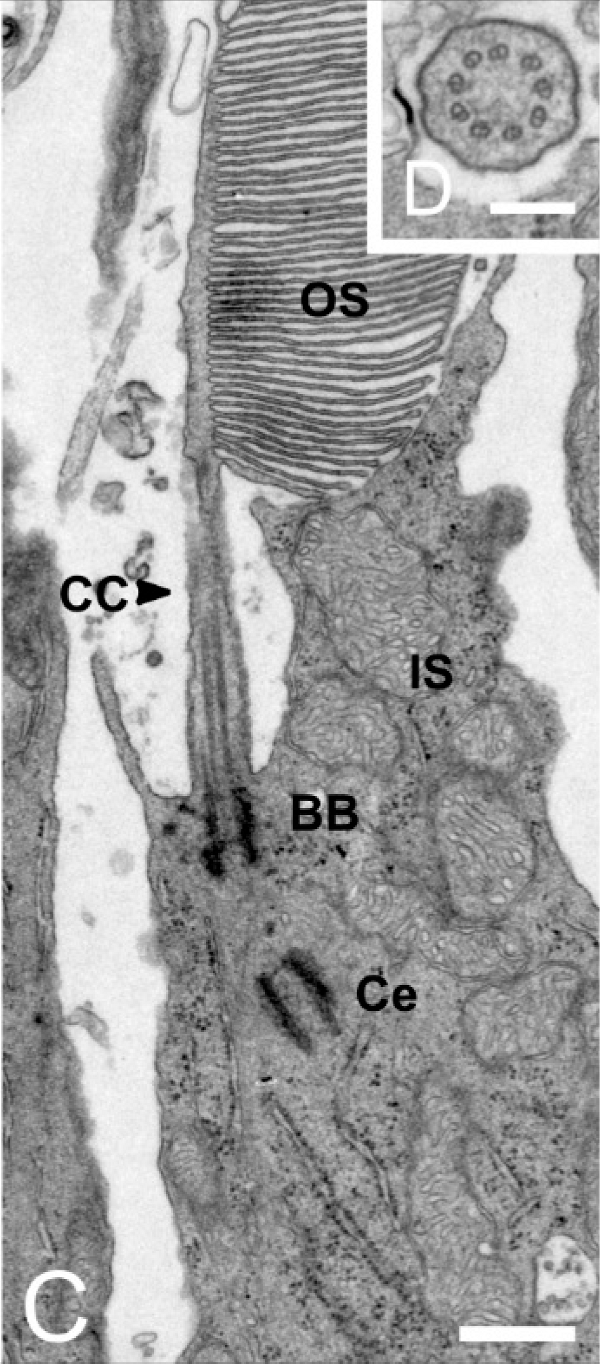
Das äußere Segment der Photorezeptorzellen ist eine umgebaute primäre Zilie21., 23.. Da die Biologen sich unter Zilien so etwas wie die Flimmerhärchen in der Lunge vorstellten, kam man erst darauf, als die Forschungsmethoden weit genug fortgeschritten waren, um sowohl den Feinbau des äußeren Segmentes als auch seine chemische Verwandtschaft mit anderen primären Zilien in Sinneszellen zu verstehen21..
Unter dem Elektronenmikroskop kann man erkennen, daß am Ansatz des äußeren Segmentes der Basalkörper der Zilie zu finden ist, bei dem es sich um zwei Zentriolen, Mutter- und Tochterzentriole handelt. An dieser setzt die Transitinszone an, die die Zilie vom Zellkörper abgrenzt. Als Verlängerung der Zentriole bildet sich die Axonemstruktur aus neun um Kreis angeordneten Paaren von Mikrotubuli aus, die das Rückrat der Zilie und Eisenbahnschiene zum Transport der in der Zilie benötigten Stoffe dient.
VB218.6.6.4
Mikrotubuli in Zilien und Flagellen
Daß man anfangs nicht darauf kam, daß es sich um eine Zilie handelt, liegt an den Membranstapeln im Äußeren Segment der Photorezeptorzellen, die bewirken daß die Zilie nicht haarartig dünn sondern fast so dick wie das innere Segment ist, das Teil des Zellkörpers ist. Diese Membranstapel setzen sich aus zusammengedrückten Bläschen, den Membranscheibchen zusammen.
2.2.3.1.3 Das äußere Segment muß als primäre Zilie mit allem beliefert werden, was dort gebraucht wird
Da es sich bei dem äußeren Segment um eine Zilie handelt, existiert dort nichts, mit dem man Proteine bauen könnte. Alles, was gebraucht wird, um das äußere Segment aufzubauen, ist als Gen in der DNA des Zellkerns codiert, wird dort transkribiert (abgelesen und als RNA übersetzt), auseinandergeschnitten und zu der richtigen RNA zusammengeklebt. Danach wird es durch das Endoplasmatische Retikulum zu den Ribosonen des rauhen Endoplasmatischen Retikulums im inneren Segment transportiert und dort in ein Protein übersetzt und mit der richtigen Adresse versehen, nämlich einer ciliaren Lokalisationssequenz (englisch: ciliary localization sequences, CLSs), und dann durch Kinesin die Mikrotubuli des Axonems entlang an ihren Bestimmungsort im äußeren Segment transportiert.22.
Wie alle Proteine wird der Sehfarbstoff Rhodopsin im Innensegment hergestellt. Vom Zellinneren wandert es die Zellmembran entlang über das Verbindungsstück in die hier noch offenen unteren Membranfalten. Diese Membranfalten werden nach und nach größer und schließen sich zu flachgedrückten Bläschen, ähnlich wie ein leerer, zusammengedrückter Luftballon, die dann mit Rhodopsin angefüllt und fertig ist und als Membranscheibchen bezeichnet werden. Dann wird die nächste Membranfalte mit Rhodopsin angefüllt und zu einem zusammengedrückten Membranscheibchen geschlossen, das dann Teil des Membranstapels ist. Mit jeder weiteren Schicht, die gebildet wird, wandern die vorher hergestellten Schichten weiter nach außen - im Bild nach oben. Verbrauchte Membranschichten werden außen von den Pigmentepithelzellen phagozytiert - also aufgefressen.
O7.19.2.2.3.2.2
Rhodopsin und Retinal - ein G-Protein gekoppelter Rezeptor, der Licht erkennen kann
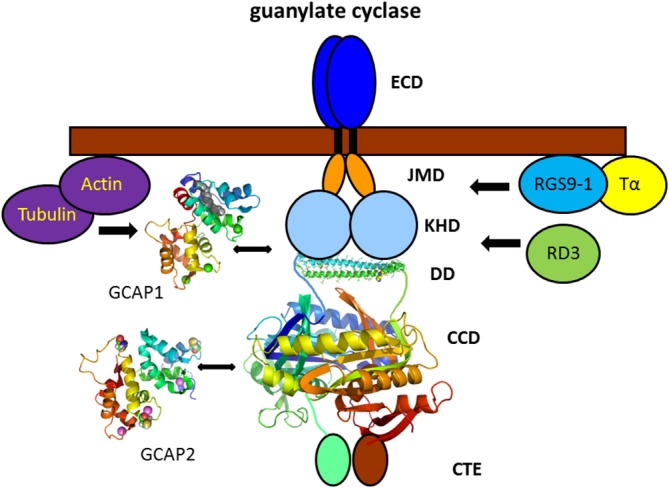
Damit die Guanylylcyclase aktivierenden Proteine GCAP1 und GCAP2 der Guanylylcyclase sagen können, daß sie den Dunkelzustand der Zelle wiederherstellen soll, müssen sie erst in der Zilie ankommen. Damit sie tatsächlich dorthin transportiert werden, müssen sie im Endoplasmatischen Retikulum an RD3 binden, das als Retinadegenerationsprotein 3 benannt wurde, weil die äußeren Segemente absterben, wenn das nicht geschieht und der Kalziumlevel dardurch im äußeren Segment so niedrig wird, daß die äußeren Segmente letztlich absterben und die betroffenen Menschen oder Tiere an Leberscher Kongenitaler Amaurose erkranken, die zu angeborener Blindheit führt39..
In folgendem beschreibe ich die einzelnen Proteine und sonstigen Moleküle in der Lichtaktivierungskette, um zu zeigen, wie sehr die Forschung dazu bereits in die Einzelheiten gegangen ist. Wer das so im Einzelnen gar nicht wissen will, kann gleich bei der Zusammenfassung weiterlesen.
O7.19.2.2.3.2.x
Zusammenfassung der Lichtaktivierungskette
Wer zu dem Ergebnis kommt: "Die wissen das so genau, so genau will ich das gar nicht wissen!" hat auch verstanden, was ich damit sagen will. Biologen wissen oft sehr viele Einzelheiten sehr genau, aber gerade wegen der vielen Einzelheiten tritt oft das "Den Wald vor lauter Bäumen nicht sehen"-Problem auf. Es besteht eben dadurch die Gefahr, daß man den Blick für die Gesamtzusammenhänge verliert, weil man sich zu sehr mit Einzelheiten befaßt.
Wenn Biologen oder Mediziner bei der Forschung Fehler machen, die nicht durch nachfolgene Forscher umgehend korrigiert werden, dann liegen die gewöhnlich nicht auf der Ebene, daß die Vorgänge, die sie beschreiben, nicht existieren würden, sondern sie liegen normalerweise darin, daß die Gesamtwirkung auf den Organismus falsch eingeschätzt wird, weil man sich zu sehr auf seinen konkreten Forschungsgegenstand konzentriert und zu wenig die unbeabsichtigten Nebenwirkungen und Quervernetzungen im System beachtet.
2.2.3.2.2 Rhodopsin und Retinal - ein G-Protein gekoppelter Rezeptor, der Licht erkennen kann
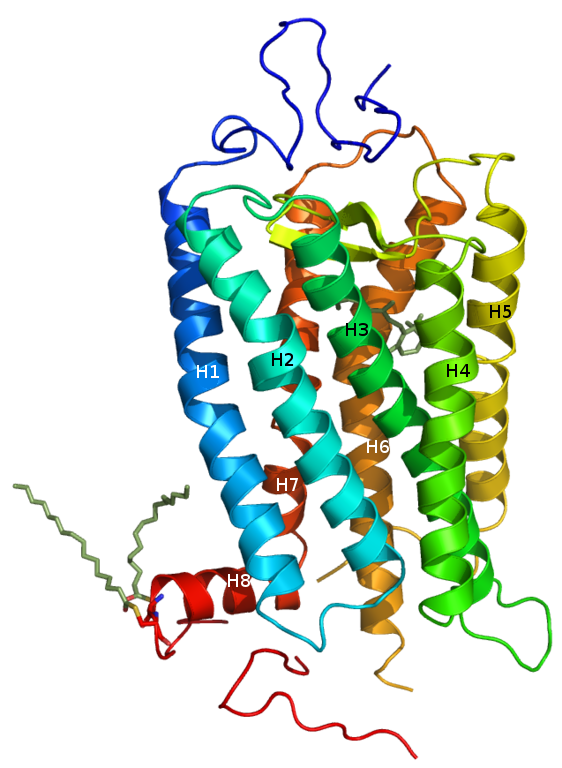
Rhodopsin, ist ein zu den G-Proteinen gekoppelten Rezeptoren zählendes Protein, das mit sieben Transmembranhelices (im Bild als H1 bis H7 durchnummeriert) durch die Zellmembran hindurchragt und das als Rezeptor der Zelle zur Lichtwahrnehmung dient. In den Stäbchen übernehmen andere Opsine - damit verwandte Sehfarbstoffe - dieselbe Funktion wie das Rhodopsin.
Inzwischen ist bekannt, wie das Rhodopsinmolekül aussieht.
Das Retinal ist im oberen Drittel zwischen H3 und H4 und vor H6 als graues Molekül zu sehen.
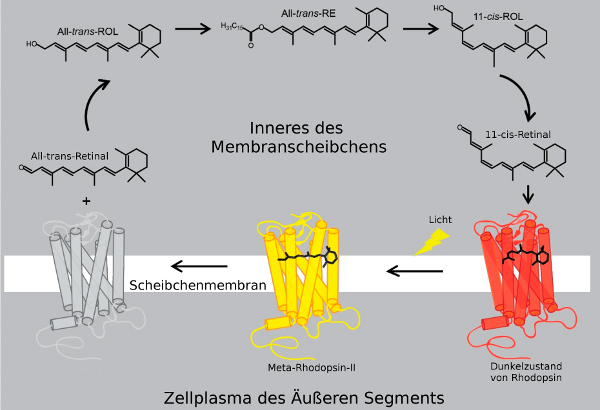
Die Alphahelix 7 (H7) von Rhodopsin ist durch eine Imin-Bindung (englisch Schiff-base bond) mit dem Retinal verbunden, daß im Dunkelzustand als 11-cis-Retinal vorliegt, das sich um einen Rest (Lys296) einer der Aminosäuren der Alphahelix 6 (H6) herumkrümmt. Alle sechs Transmembranhelices sind in diesem Zustand eng miteinander verbunden. Wenn Licht auf das Retinal fällt, streckt sich das Molekül und wandelt sich so in All-trans-Retinal um. In dieser Form drückt es die Helices an der Außenseite der Membran, wo sich das Retinal befindet, auseinander. H1, H2, H3 und H4 sind durch ihre Reste Reißverschlußartig so eng miteinander verbunden, daß sie im Dunkelzustand des Rezeptors in derselben Stellung sind wie bei Licht, H5, H6, H7 bewegen sich, wenn Licht auf das Molekül fällt, um einige Bindungen herum, die wie ein Scharnier wirken, so daß im inneren Zellplasma (unten) die Bindungsstelle für das G-Protein Transducin (Gt) frei wird. Dieser aktivierte Zustand des Rhodopsins heißt Meta-Rhodopsin-II.
25., 28., 29.
Ob das Rhodopsin im Dunkelzustand ist oder bereits durch Licht zu Meta-Rhodopsin-II aktivert ist kann man auch direkt sehen. Während Rhodopsin im Dunkelzustand der Retina eine rote Farbe verleiht - daher der Name Sehpurpur - kann es im lichtaktivierten Zustand kein Licht mehr absorbieren. Daher wirkt die Retina nach Bestrahlung mit Licht viel heller40..
Zeichnung nach einem Mikroskopischen Bild aus dem Artikel, in dem Franz Boll beschrieben hat, wie sich Rhodopsin - der Sehpurpur - aufhellt, wenn man die Retina mit Licht bestrahlt. Er hat diese Experimente mit Fröschen gemacht. Die roten Kreise sind Stäbchen, andersfarbigen Kreise stellen Zäpfchen dar.40.
Transducin zählt zu den G-Proteinen, die den G-Proteinen gekoppelten Rezeptoren den Namen gegeben haben. Es selbst wiederum hat seinen Namen von Guanosin-Di-Phosphat (GDP) und Guanosin-Tri-Phosphat (GTP), seinem Bindungspartner.
VB218.7.1.2
Guanin, Gunanosin und seine Phosphate GDP und GTP
Das zu den G-Proteinen gehörende Transducin in Bändermodelldarstellung. Es handelt sich um ein heterotrimeres G-Protein, hetero heißt verschieden, tri drei und die Nachsilbe -meres macht daraus dreiteilig. Ein heterotrimeres G-Protein ist also ein G-Protein aus drei verschiedenen Teilen. Blau ist die αt/αi-Untereinheit dargestellt, an die ADP oder ATP gebunden wird. Rot und grün sind β- und γ-Untereinheit dargestellt.
Wenn das Rhodopsin im Dunkelzustand ist, kann das G-Protein Transducin nicht daran binden, da die Bindungsstelle erst im aktivierten Zustand Meta-Rhodopsin-II frei wird. Das Transducin befindet sich daher auf Zellplasma-Seite der Membran (im oberen Bild unten) und wartet im inaktiven Zustand, in dem das energieärmere GDP daran gebunden ist, darauf daß das Rhodopsin durch Licht angeregt wird und das darin gebundene Retinal sich streckt, dafür sorgt, daß das Rhodopsin zu Meta-Rhodopsin-II wird und die Bindungsstelle frei wird. Dann bindet sich das Transducin an das Meta-Rhodopsin-II und das GDP wird durch GTP ersetzt, wodurch Transducin in zwei Teile zerfällt die β- und Gamma Untereinheit und die α-Untereinheit. Die α-Untereinheit von Transducin trägt jetzt das ATP und ist daher im aktiven Zustand, in dem sie das Signal zur Phosphodiesterase weitertragen kann.
Im Dunkelzustand des Stäbchens ist im Zellplasma des äußeren Segments reichlich cyclisches Guanosinmonophosphat (cGMP) vorhanden und das Membranpotential der Zilienmembran ist relativ gering, da die cGMP-abhängige Kationenkanäle (engl. cGMP-gated cation channels) deshalb offen sind. Dieser Zustand wird durch Guanylylcyclasen aufrecht erhalten.
Die Phosphodiesterase 6 (PDE6) wandelt cyclisches Guanosinmonophosphat (cGMP) in Guanosinmonophosphat (GMP) um, was zur Schließung der cGMP-abhängige Kationenkanäle (engl. cGMP-gated cation channels) und zu einer Hyperpolarisation der Zellmembran führt.
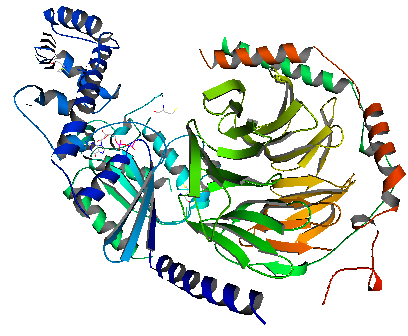
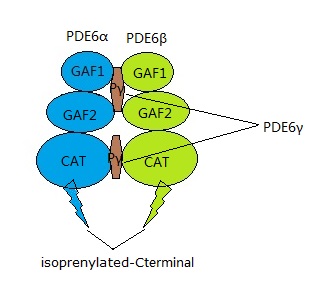
Die Phosphodiesterase 6 (PDE6) besteht im Stäbchen aus drei Typen von Untereinheiten. PDE6α und PDE6β sind einander sehr ähnlich und sind durch die Untereinheit PDE6γ miteinander verbunden. PDE6γ besetzt im inaktiven Zustand der Phosphodiesterase 6 genau an die Stellen der anderen beiden Untereinheiten PDE6α und PDE6β, die für die Phosphodiesterase-Funktion zuständig sind. Erst wenn die durch GTP aktivierte α-Untereinheit von Transducin andockt, löst sich PDE6γ dort, die Phosphodiesterase kann aktiv werden und cyclisches Guanosinmonophosphat (cGMP) in Guanosinmonophosphat (GMP) umwandeln. Die Phosphodiesterase 6 (PDE6) unterscheidet sich von der mit ihr am nächsten verwandten Phosphodiesterase 5 (PDE5), die in der glatten Muskulatur der Blutgefäße des Penisschwellkörpers und der Lunge sowie in Thrombozyten auftritt, vor allem dadurch, daß sie ungefähr tausend mal so leistungsfähig ist, wenn sie aktiviert wird, da im Stäbchen eine ausgesprochen hohe Leistung benötigt wird.30.
2.2.3.2.5 RGS9-1 (Regulator of G Protein Signaling 9, splice variant 1)
RGS9-1 (Regulator of G Protein Signaling 9, splice variant 1) - deaktiviert die α-Untereinheit von Transducin (Tα) wieder, so daß der Dunkelzustand nach Ablauf der Signalkette der Lichtaktivierung wiederhergestellt werden kann. Man ist ewig lange geblendet, nachdem man einen Lichtblitz abbekommen hat, wenn RGS9-1 fehlt37., weil Transducin dann einfach weiterarbeitet.
Die cGMP-abhängigen Kationenkanäle (engl. cGMP-gated cation channels) gibt es in den Stäbchen und Zäpchen des Auges. Ähnliche Kanäle, die aber auch auf cAMP reagieren gibt es in Geruchssinneszellen, chemosensitiven Zilien und der Zirbeldrüse (oder Epiphyse, Corpus pineale, englisch Pineal body).35.
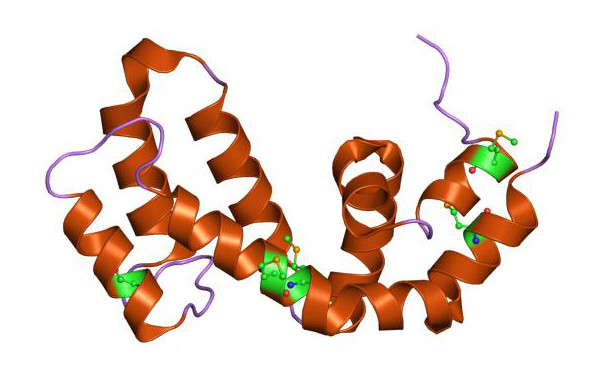
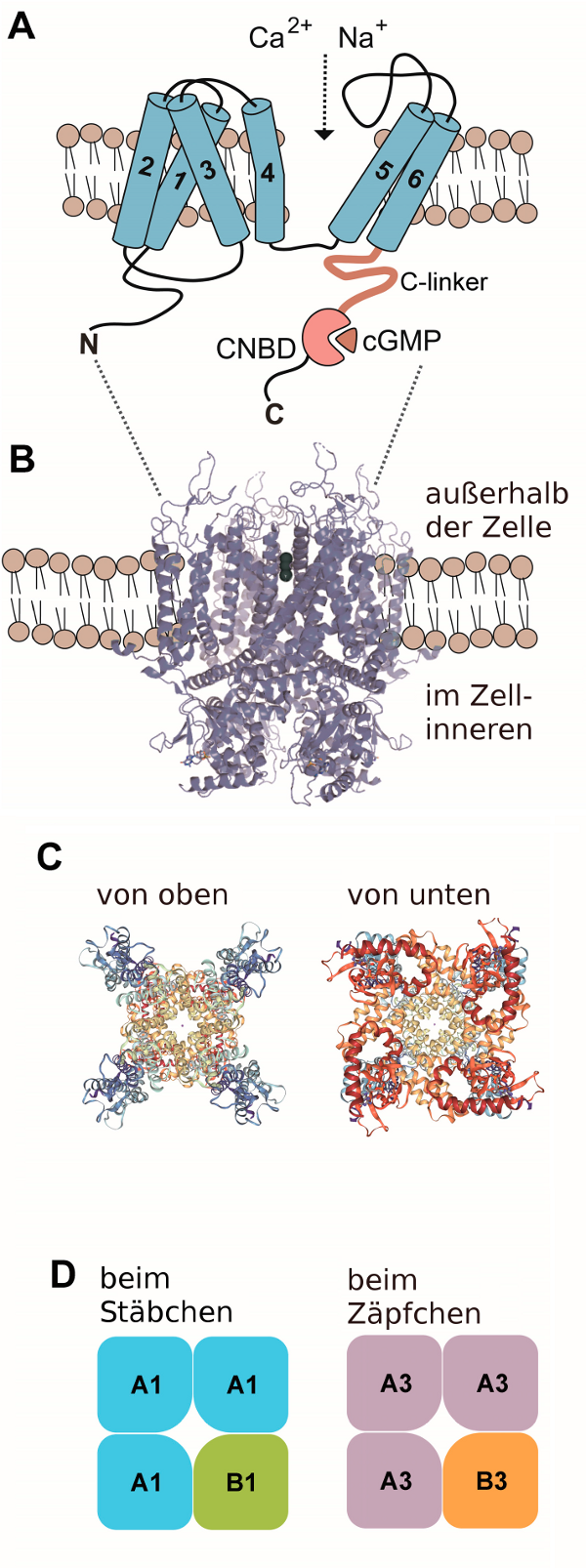
(A) Die Anordnung der Alpha-Helices 1–6 einer einzelnen Untereinheit des Kanals, C, carboxy-terminus; CNBD, cyclic nucleotide binding domain; N, amino-terminus.
(B) Bändermodell des Kanals von Caenorhabditis elegans und seine Lage innerhalb der Membran.
(C) Anblick von oben und unten
(D) Zusammensetzung aus unterschiedlichen Untereinheiten beim Stäbchen und Zäpfchen. A1, CNGA1; A3, CNGA3; B1, CNGB1; B3, CNGB3.
Nachdem RGS9-1 die α-Untereinheit von Transducin (Tα) wieder deaktiviert hat, stellt die Guanylylcyclase cyclisches Guanosinmonophosphat (cGMP) aus Guanosintriphosphat (GTP) her, indem sie zwei Phosphatgruppen abspaltet und den Rest zu einem Ring zusammenfügt.
Guanylyl-Cyclase aktivierenden Proteine GCAP1 und GCAP2 regeln die Aktivität der Guanylylcyclase bei niedriger Ca2+-Ionenkonzentration im Plasma der Zilie hoch und sorgen so dafür, daß die Guanylylcyclase den Dunkelzustand der Zelle wieder herstellt.33.
Im Dunkelzustand des Stäbchens ist im Zellplasma des äußeren Segments reichlich cyclisches Guanosinmonophosphat (cGMP) vorhanden und das Membranpotential der Zilienmembran ist relativ gering, da die cGMP-abhängige Kationenkanäle (engl. cGMP-gated cation channels) deshalb offen sind. Dieser Zustand wird durch Guanylylcyclasen aufrecht erhalten.
Darstellung der Guanylylcyclase des Photorezeptos der Wirbeltiere und der Proteine, die mit ihr zusammenarbeiten. Die Proteinbestandteile, deren genaue Struktur 2015 bekannt war, sind durch Bänderdiagramme, die anderen nur schematisch dargestellt.
Die Bestandteile der Guanylylcyclase, die hier "guanylate cyclase" genannt ist, sind:
ECD, extracellular domain - der Proteinbestandteil, der sich im Inneren des Membranscheibchens befindet
TM, die Transmembrandomäne - der Teil, der die Scheibchenmembran durchdringt
JMD, juxtamembrane domain - der Teil der der Membran anliegt
KHD, kinase homology domain - nach der ähnlichkeit mit Kinasen benannt
DD, dimerization domain
CCD, cyclase catalytic domain - der Teil der Cyklase der das cyclische Guanosinmonophosphat (cGMP) herstellt
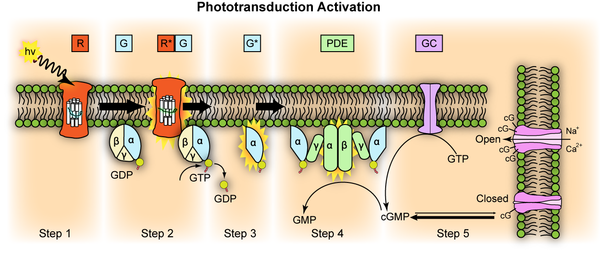
Repräsentation der molekularen Schritte der Lichtaktivierung. Die waagerecht dargestellte Membran ist die Scheibchenmembran, die senkrechte Membran ist die Zilienmembran.
Das eintreffende Photon (hv) wird absorbiert und aktiviert das Rhodopsin durch eine Konformationsänderung in der Scheibenmembran zu Meta-II-Rhodopsin (R*).
Als nächstes hat Meta-II-Rhodopsin wiederholten Kontakt zu Transducin-Molekülen.
Dies katalyisiert die Aktivierung zu G* durch die Freisetzung von gebundenem GDP und bindet dann freies GTP. Die Alpha- und Gamma Untereinheiten von G* binden an die hemmende Gamma Untereinheit der Phosphodiesterase (PDE), was seine Alpha- und Beta Untereinheiten aktiviert.
Aktiviertes PDE hydrolysiert cGMP zu GMP. Reduzierte Mengen von freiem cGMP führen dazu, dass die Ionenkanäle in der Zellmembran geschlossen werden und weiteren Einstrom von Na+ und Ca2+ verhindern. Die Photorezeptorzelle hyperpolarisiert.
Erst wenn die Phosphodiesterase (PDE) nicht mehr ständig neu aktiviert wird, weil das Opsin wieder in den Dunkelzustand zurückgekehrt ist, kann die Guanylyl-Cyclase (GC) die Oberhand gewinnen und genug cGMP prouzieren, welches als second messenger, als zweiter Botenstoff fungiert und den Dunkelzustand wiederherstellt, so daß die Ionenkanäle wieder öffnen.
2.2.4.1 Die Evolution der Lichtsinneszellen und ihrer Farbstoffe
Das Rhodopsin gehört zu den Opsinen, die von verschiedensten Lebewesen, zur Lichtwahrnehmung verwendet werden. Die Ahnen der Säugetiere, die Therapsiden, waren Tetrachromaten. Sie hatten vier Sorten Zäpfchen mit jeweils unterschiedlichen Sehfarbstoffen, wie es heute beispielsweise die meisten Vögel und Reptilien auch haben. Wir Menschen sind Trichromaten. Die meisten Säugetiere, die keine Beuteltiere sind, sind ähnlich ausgestattet, wie rot-grün-farbenblinde Menschen. Bei den höheren Primaten hat sich der ursprünglich für rot vorgesehene Farbstoff genetisch verdoppelt und etwas verändert, so daß wir jetzt wieder Trichromaten sind. Erklärt wird der Verlust der Opsine aus den Zäpfchen und divereser anderer Opsine, die auch Licht wahrnehmen können, auch wenn sie nicht dem sehen im engeren Sinne dienen, dadurch, daß Säugetiere ursprünglich nachtaktiv waren und daher mehr auf hohe Lichtempfindlichkeit der Sinneszellen und weniger auf Farbtüchtigkeit angewiesen waren.41., 42.
VB230.1.1.2
Die Opsine der Wirbeltiere
Nicht bei allen Menschen ist die Farbwahrnehmung gleich. Es gibt einige Menschen die rot-grün-farbenblind sind und damit noch die Farbwahrnehmung unserer dichromatisch veranlagten Ahnen haben.
Es gibt bei Menschen eine Mutation des Opsins für Rot, die bei mischerbigen Frauen, die in unterschiedlichen Zäpfchen unterschiedliche X-Chromosomen desaktiviert haben, dazu führt, daß vier verschiedene Opsine vorhanden sind. Solche Frauen können mehr unterschiedliche Farben wahrnehmen als normale Menschen, da sie Tetrachromaten sind. Sie konnten in einem Farbspektrum gewöhnlich 10 Farben unterscheiden, Trichromaten konnten durchschnittlich 7,3 Farben unterscheiden und Rot-Grün-Farbenblinde 5,3 Farben.63., 64. Daß die Tetrachromaten immer Frauen sind und die rot-grün Farbenblinden meist Männer sind, ist offensichtlich die Erklärung dafür, warum Männer in den verschiedensten Kulturen weltweit weniger Farben unterscheiden als Frauen.65.
2.2.4.2 Die Zäpchen
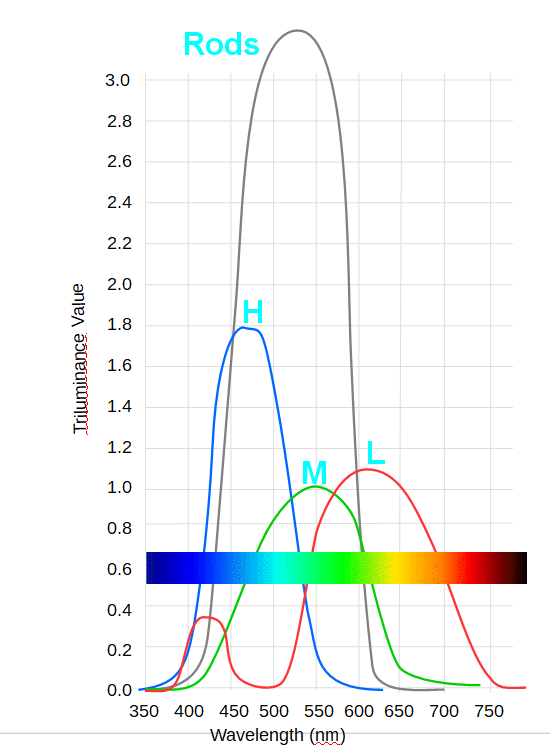
Da wir von nachtaktiven Tieren abstammen, haben wir wie die meisten Säugetiere sehr viele Stäbchen in der Retina und weitaus weniger Zäpfchen, denn die Stäbchen sind sehr viel lichtempfindlicher als die Zäpfchen.
Stärke der Reaktion der Stäbchen ("rods" schwarz) und verschiedenen Zäpfchen auf Licht. Deutlich erkennbar ist das die Stäbchen viel stärker auf Licht reagieren als die Zäpchen.
Da das Zäpfchen für blau lichtempfindlicher ist als die für grün, sehen wir helle Stellen eines Gegenstandes etwas gelblicher und die Stellen, die im Schatten liegen, wirken bläulich. Wenn man ein Bild malt, wirkt es daher echter, wenn man an den hellen Stellen etwas gelb unter die Farbe mischt und an den dunklen etwas blau.
Daß die Zäpfchen für Rot zwei Wahrnehmungsmaxima haben, von denen einer am Rand des blauben bereiches liegt, führt dazu, daß wir die Farbe violett wahrnehmen, die eigentlich so nicht existiert und die farben intuitiv in einem Farbkreis anordnen.
O7.D5.3.1
Der Irrtum im Farbkreis
2.2.4.3 Verteilung von Stäbchen und Zäpchen im Auge
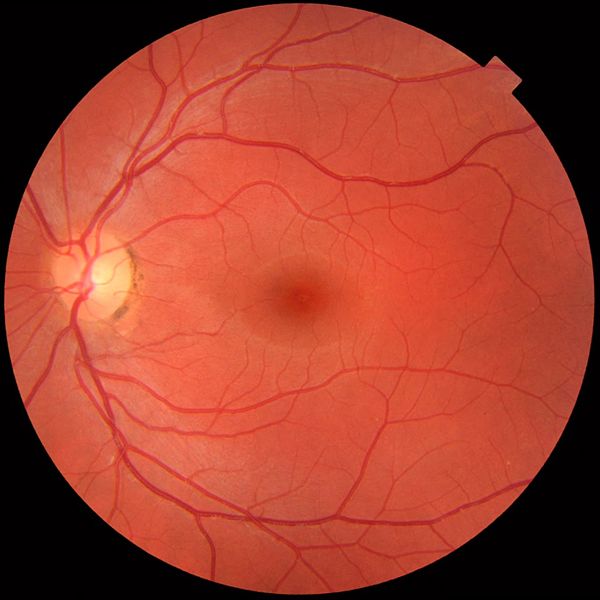
Wenn man durch die Augenlinse hindurch mit einer Spezialkamera das innere des Auges fotographiert, entsteht folgendes Bild.
Bildquelle:
Fundus vom linken Auge. Der hell Fleck links, ist die Sehnervenpapille, der blinde Fleck des Auges, wo die Adern und Nerven den Augapfel verlassen und wo deshalb kein Platz für Stäbchen und Zäpfchen ist. Er liegt zur Nase hin. Die rot verzweigten Arterien, durch die das Blut ins Auge transportiert wird sind heller und etwas dünner als die entsprechenden Venen, durch die das Blut das Auge verläßt. Die Nerven sind als dünne helle Striche zu erkennen, wenn man genau hinsieht.
Der dunkle Fleck in der Mitte ist die Fovea centralis, die Stelle der Netzhaut, mit der wir am schärfsten sehen.
Eine Linse bricht - wenn man keine Gegenmaßnahmen ergreift - Licht unterschiedlicher Wellenlängen unterschiedlich stark. In optischen Geräten wird das ausgeglichen, indem Linsensysteme aus unterschiedlichen Materialien verwendet werden. In der Augenlinse selbst ist dieser Fehler nicht korrigiert. In der Fovea centralis, der Stelle schärfsten sehens wird der Fehler jedoch vermieden, indem es dort nur Zäpfchen für rot und grün gibt, so daß die unterschiedlichen Brechungsindizes für unterschiedliche Farben dort nicht ins Gewicht fallen, da die Größe der hier einzeln angeschlossenen Zäpfchen die Schärfe des Sehens begrenzt. Weiter am Rand des Gesichtsfeldes, wo es auch blaue Zäpfchen gibt, sind mehrere Zäpfchen für das Sehen miteinander verschaltet und diese Verschaltung begrenzt die Auflösung des Auges, so daß auch hier der Brechungsfehler nicht ins Gewicht fällt.54.
Zäpfchenmosaik in der Fovea centralis, links bei einer normalsichtigen, rechts bei einer rot-grün farbenblinden Person, bei der die rot-empfindlichen Zäpfchen fehlen. Man beachte daß die Zäpfchen für blaues Licht in der Mitte fehlen.
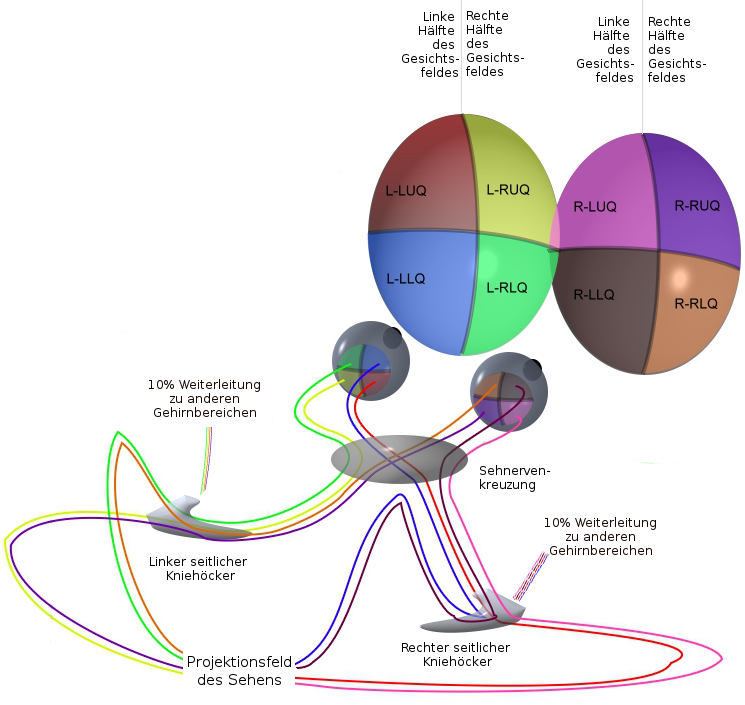
Die Nervenbahnen vom Auge laufen zur Sehnervenkreuzung. Da wechselt ein Teil der Fasern zur gegenüberliegenden Seite. Von dort aus laufen sie zum seitlichen Kniehöcker im Metathalamus des Zwischenhirns, wo sie auf andere Nerven umgeschaltet und zur Projektionsfeld des Sehens weitergeleitet werden. Während 90 Prozent der von dort weitergehenden Nervenbahnen zum Projektionsfeld des Sehens führen, gehen die anderen 10% in andere Bereiche des Gehirns, wo sie unbewußt weiterverarbeitet werden. Diese 10% anderen Nerven sind für das blinde Sehen verantwortlich.2.
VB160.
Blindes sehen
Im Projektionsfeld des Sehens bildet die Netzhäute Punkt für Punkt ab. Allerdings nicht einzeln, sondern es ist jeweils so, daß ein Streifen, der Nervenfasern vom rechten Auge erhält, neben einem Streifen liegt, der Nervenfasern vom entsprechenden Teil des Gesichtfeldes vom linken Auge erhält. Insgesamt ergibt sich ein Muster ähnlich einem Fingerabdruck.61.
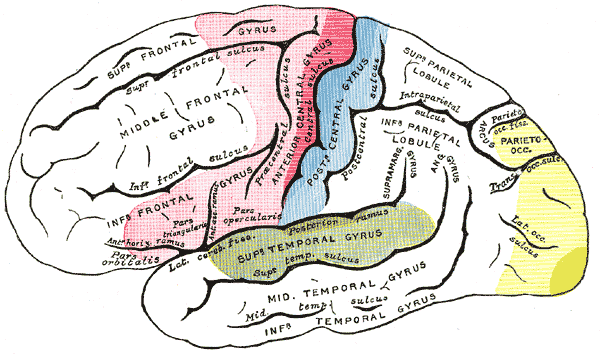
Gehirn von außen:
Das Sehzentrum ist gelb markiert. Der dunkelgelbe Bereich ist das Projektionsfeld des Sehens, das auch primäre Sehrinde genannt wird. Der hellgelbe Bereich ist das Assoziationsfeld des Sehens, das dazu dient, das Gesehene zu verstehen.
Das Assoziationsfeld des Sehens liegt ebenfalls in dem Großhirnteil am Hinterkopf, wo auch die primäre Sehrinde liegt. Es umgibt die primäre Sehrinde auf allen Seiten. Dieser Gehirnteil dient dazu das Gesehene zu erkennen. Das geschieht einerseits auf der bildlichen Ebene, indem wir erkennen, daß ein Pferd ein Pferd ist.
O7.4.6
Störungen beim Erkennen des Gesehenen: Der Gestalterkennungsfilter
Wir ordnen in einem Teil dieses Bereiches aber auch beispielsweise geschriebene Wörter ihrer Bedeutung zu.
Umgekehrt kann etwas, das sich nachträglich als bestätigbare feinstoffliche Erfahrung erweist, in der subjektiven Wahrnehmung von einem sehr vagen Eindruck bis hin zu einem fast realistischen sehen, das man mit einer irdischen Wahrnehmung verwechseln kann, rangieren.
Aus der subjektiven Wahrnehmungsqualität kann man daher nicht sicher schießen ob etwas eine irdische Wahrnehmung, eine Halluzination oder ein Traum, oder aber eine feinstoffliche Wahrnehmung eines realen Sachverhaltes ist.
Nachdem ich mich mit diversen unterschiedlichen Hellsichtigen unterhalten habe und einiges darüber gelesen habe, kann ich sagen: Wenn jemand häufig feinstoffliche Wahrnehmungen hat, auf die er sich verläßt, weil sich das bisher als eine gute Taktik erwiesen hat, kann es aber sein, daß er diese Wahrnehmungen an irgendeinem Merkmal erkennen kann. Es mag daran liegen, daß es sich wahr anfühlt, es kann eine besonders realistische Wahrnehmungsqualität sein, es kann aber auch sehr vage und kaum glaubhaft erscheinen und die Person weiß trotzdem daß es Kommunikation ist, die er als solche erkennt. Anders ausgedrückt: Während es kein Kriterium gibt, das jeder Mensch gleichermaßen als Unterscheidungsmerkmal zwischen feinstofflicher Wahrnehung und unbedeutenden Traumbildern nehmen kann, haben diverse Menschen jedoch ein internes Signal, mit dem sie sich selbst mitteilen, ob etwas Fantasie, Traum, irdische oder feinstoffliche Wahrnehmung ist. Dies Signal ist aber von Mensch zu Mensch verschieden.
Trotzdem - wenn man beweisen will, daß es so etwas wie feinstoffliche Wahrnehmung überhaupt gibt, ist Wahrnehmungsqualität offensichtlich in keiner Hinsicht beweiskräftig, da Dinge die wir real vor Augen haben, Halluzinationen, Illusionen und feinstoffliche Wahrnehmungen dieselben Wahrnehmungsqualitäten haben können.
2.2.8 Es gibt keine Spiegelneuronen, sondern nur ein neuronales Spiegeln
Spiegelneuronen ist ein Begriff der in der Welt herumschwirrt und den ich ehrlich gesagt sehr unpassend finde, denn das System das so genannt wird, hat diverse Funktionen, vor allem die, eine Handlung mit ihrem Sinn zu verbinden, beispielsweise ein Sinn wie "nach etwas greifen" oder "etwas wegwerfen". Für diesen Zweck wird es bei der Planung der eigenen Handlungen verwendet, beim einfachen Nachdenken über Handlungsmöglichkeiten, aber auch um fremde Handlungen zu interpretieren. Wir verstehen fremde Handlungen also, indem wir die Bewegungen und ihren Sinn im eigenen Gehirn simulieren und nur die nachgeschalteten Schritte weglassen, indem wir die Arbeit der Neuronen mit der Funktion der konkreten Muskelbewegung, beispielsweise "Bewege den Beugemuskel für den rechten kleinen Finger" weglassen. Ich glaube daß die italienischen Autoren, die ja Biologen und keine berufsmäßigen Übersetzer sind
62.
VB253.
Es gibt keine Spiegelneuronen, sondern nur ein neuronales Spiegeln
Bei Ringelwürmern (Annelida)48., Insekten55., Fischen (Teleostei)44., Amphibien51., Reptilien49., Vögeln (Aves)46., 47., 50., Säugetieren (Mammalia)45., 50. und auch beim Menschen selbst wurden mitten im Gehirn Sehfarbstoffe nachgewiesen, die allerdings nicht direkt zum sehen verwendet werden. Da fragt man sich natürlich, was Lichtsinneszellen im Gehirn suchen, von dem zumindest ich früher angenommen habe, daß es dort mitten im Schädel dunkel ist.
Ich erzählte einer Freundin davon und sie meinte:
"Vielleicht gibt es da so etwas wie Glasfaserkabel, die das Licht ins Gehirn leiten."
"Hmm, irgendsoetwas haben sie tatsächlich nachgewiesen." antwortete ich und erzählte von einen wissenschaftlichen Artikel, wo sie den menschlichen Kopf - gut abgeschirmt - von einer Seite beleuchtet und dann nachgeschaut haben, ob auf der anderen Seite etwas davon wieder herauskommt. Es war auf der anderen Seite tatsächlich Licht meßbar56.. Auf den Gedanken, daß es im Körper Lichtwellenleiter geben muß, die ähnlich wie Glasfaserkabel funktionieren, sind auch Wissenschaftler schon gekommen. Sourabh Kumar et Al. haben ausgerechnet, daß die Axone der Nervenzellen zu diesem Zweck geeignet sein müßten58.. Shelli R. Joye stellt diese Überlegung auch an und schlägt einerseits die Blutkapillaren vor, andererseits die Mikrotubuli59.. Die Idee mit den Blutkapillaren hat mich nicht überzeugt, zu den Mikrotubuli gibt es dagegen Belege, daß sie tatsächlich eine Funktion im Bewußtsein haben die durch ihre Funktion als Eisenbahnschienen der Zelle und als Teil des Zellskellettes nicht so ganz zu erklären sind.
VB209.2.2
Was verbindet die Mikrotubuli mit dem Bewußtsein
Ein anderer Gedanke, ist daß das Vorhandensein von Opsinen im Gehirn möglicherweise etwas mit Biophotonen zu tun hat, die ja teilweise auch im Bereich des sichtbaren Lichtes liegen60. und daß diese Opsine daher möglicherweise biophotonische Signale in chemische Signale übersetzen.
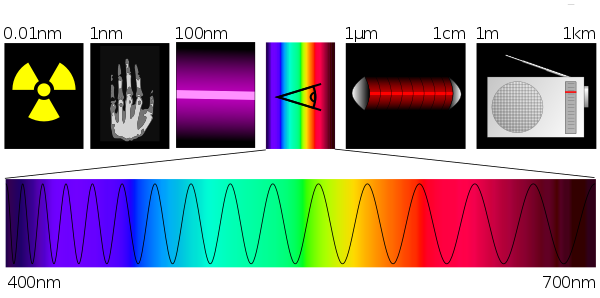
Das sichtbare Licht ist nur ein kleiner Ausschnitt aus dem Gesamtspektrum der elektromagnetischen Strahlung.
Bei hoher Schwerkraft von über +9g werden zuerst die am weitesten von der versorgenden Ader entfernten Ränder der Netzhaut nicht ausreichend versorgt, es entsteht deshalb eine Tunnelvision. Dies wird von Whinnery (1997) als ein gemeinsamer Bestandteil der G-LOC-Erfahrungen mit den Nahtodeserlebnissen beschrieben. Tatsächlich entspricht die Tunnelerfahrung in der Nahtoderfahrung nicht diesem Bild, da neben dem klassischen Tunnel mit dem Licht am Ende auch jede Art anderer Tunnel auftreten kann, die Tunnelerfahrung nicht immer am Anfang sondern häufig auch zwischen Außerkörperlicher und Jenseitserfahrung auftritt und das Auftreten der Tunnelerfahrung nicht von der Art der Lebensgefahr abhängt, aber gelegentlich durch völlig andere Symboliken ersetzt zu sein scheint.
6., 7. S.238, 8.
O7.32.2
Tunnelblick im Vergleich zur Tunnelerfahrung
Warum Bereiche der Großhirnrinde bei der Verarbeitung der Nahtodeserfahrungen beteiligt sein müssen
Da im Nahtodeserlebnis die Identität/das Ich erhalten, Wahrnehmung kategorisiert, episodische Langzeit-Gedächtnisfunktionen aktiviert, ethisch bewertet und ein personaler und extrapersonaler Raum und damit eine kognitive stabile Umwelt geschaffen werden, ist eine Beteiligung des parietalen, temporalen und (prä)frontalen (Assoziations-)Cortex anzunehmen. Die schon während des Nahtodeserlebnisses erfolgende Bewertung der Bedeutung des Nahtodeserlebnisses benötigt beispielsweise neben dem Cortex das Raphe- und das Locus-coeruleus-, das Gedächtnis- und das limbische System, den präfrontalen Cortex, das cholinerge System des basalen Vorderhirns sowie die thalamischen Kerne.
1.6, 1.8,
1.11
Die im Nahtodeserlebnis erfolgende Aktivierung des semantischen Gedächtnisses und der Objektidentifizierung, aber auch der Luzidtraumcharakter des NDEs sprechen ebenfalls für eine Aktivierung großer Bereiche corticaler Strukturen. Auch die Musikwahrnehmung und deren Bewertung im Nahtodeserlebnis benötigen den Gyrus temporalis posterior superior der rechten und das Planum temporale des linken Temporallappens. Selbst das Träumen wird primär im Frontalhirn generiert, da ein Ausfall des mittleren und unteren Frontallappens (in beiden Hirnhälften) zu einem sofortigen Traumausfall führt.
1.6, 1.11
Funktionen des Schläfenlappens, die seine Beteiligung an der Nahtodeserfahrung nahelegen
Morse und Kollegen haben ein Modell vorgeschlagen, das auf dem Neurotransmitter Serotonin basiert und die Schritte bis zur Auslösung von Nahtod-Erfahrungen erklären soll. Dabei wird dem Schläfenlappen, auch Temporallappen genannt, eine zentrale Bedeutung beigemessen. Das auch als Schläfenlappen bekannte Großhirnareal beherbergt mit Amygdala und Hippocampus zwei wichtige Bestandteile des limbischen Systems, bei denen vieles darauf hindeutet, daß sie an der Nahtodeserfahrung beteiligt sind.
1.6, 3.3 S.243
Im Koma zeigt eine erhöhte P300-Amplitude im EEG internes Orientierungsverhalten und einen Gedächtnisabgleich an. Das führt dazu daß das Acetylcholin-übertragende Transmittersystem in der Formatio reticularis eine Überwachheit hervorruft, die vor allem das untere vordere Vorderhirn (Nucleus basalis) aktiviert. Durch diese Aktivitätssteigerung im Gehirn kommt es bei künstlich ausgelösten Hypoxien zur dissoziierten Enthemmung und damit Aktivierung bestimmeter Teile der Gehirnrinde und darunterliegebereiche Bereiche, insbesondere Teile des Limbischen Systems im Bereich der Schläfen, die weniger empfindlich auf Sauerstoffmangel und zu viel Kohlendioxid reagieren.
1.6, 1.9
O7.74
Sauerstoffmangel als denkbare Ursache Nahtodeserfahrung
Verschiedene Nahtodeserfahrungs-Qualitäten wie traumhafte Zustände, lebhafte sicht- oder hörbare Halluzinationen, das Gefühl, das erlebte sei persönlich sehr bedeutsam, starke Gefühle, das spüren von Schwingungen, Schwebegefühle, Außerkörperliche Erlebnisse, plötzliche Erkenntnisse, mystische und religiöse Erlebnisse, plötzlichen Auftauchen längst vergessener Erinnerungen i. S. von Lebensfilm-Bruchstücken und Traumsequenzen können durch künstliche oder spontane elektrische Entladungen der Schläfenlappen hevorgerufen werden. Sie beziehen wie auch die Nahtodeserlebnisse das ganze Selbst-Gefühl der Person mit ein. Bei ihnen wird eine kosmische Bedeutsamkeit empfunden und sie haben einen starken Einfluß auf das spätere Verhalten, Erleben und das Glaubenssystem der durch diese Erfahrungen betroffenen Personen. Bei künstlicher elektrischer Reizung des Schläfenlappens und durch epileptische Anfällen können Tunnelerlebnisse ausgelöst werden.
1.1, 1.6,
1.8, 1.10,
1.11,
3.3 S.44
Dissoziative Erfahrungen - zu denen Nahtodeserlebnisse und Außerkörperliche Erlebnisse zählen - führen zu einer gesteigerten oder auffälligen Aktivität des Temporallappens. Patienten mit PTLE-(Possible Temporal Lobe Epilepsy)Symptomen berichten dementsprechend vermehrt von subjektiven paranormalen und kosmisch-mystischen Erlebnissen sowie von Außerkörperlichen Erlebnissen.
1.6, 1.9
Dementsprechend entwickeln Patienten mit einer Schläfenlappen-Epilepsie unter LSD-Einfluß deutlich mehr Veränderungen der Wahrnehmung, wenn der Krampffokus in der rechten (anstatt in der linken) Gehirnhälfte sitzt und es kommt bei Schläfenlappenepilepsie häufiger zu optischen Halluzinationen, wenn der Herd in der nicht-dominanten - das ist meistens die rechte - Gehirnhälfte liegt.
1.6, 1.9
Bezug zur Drogenerfahrung: Die NMDA-Rezeptoren finden sich im Schläfenlappen
LSD wirkt über eine Herabsetzung der Serotonin-Konzentration, deren Abfall die LSD-Wirkung fördert und deren Anstieg sie hemmt. Dabei soll LSD die inhibitorische Wirkung des Nucleus raphe (dorsalis) aufheben, der wiederum mit seinen Axonen das Frontalhirn und den Temporallappen hemmt, so daß LSD Entladungen im Temporallappen bewirken kann, die zu Nahtodeserlebnisähnlichen Erfahrungen führen. Die gleiche Wirkung haben experimentell alle Halluzinogene auf den Nucleus raphe medialis, dessen Nervenfasern das limbische Vorderhirn beeinflussen.
1.6
Auch die Nahtodeserlebnis-Auswirkungen ließen sich so erklären: Eine Ausschaltung der hemmenden serotonergen Einflüsse auf das temporolimbische System soll über einen kindling-Effekt dessen Erregbarkeit langfristig erhöhen. Insbesondere die Ausschaltung der Serotonin-Freisetzung in den Pypramidenzellen des Hippocampus soll für religiöse Erlebnisse und Ekstase-Erfahrungen wichtig sein. Eine Aufhebung der serotonergen Hemmung der CA3-Zellen des Hippocampus wird auch für Amphetamine, Kokain, Halluzinogene und Ketamin angenommen, die alle NDE-Elemente produzieren können.
1.6
Serotonerge Fasern des Nucleus raphe dorsalis stehen mit der Griseum centrale in Verbindung. Das ist der Ort mit höchster Endorphin-Konzentrationen im Gehirn. Serotonin spielt außerdem eine Rolle bei der Reduktion der Schmerzwahrnehmung unter Streß. Eine Erhöhung des Co2-Gehaltes im Blut kann zu Sauerstoffmangel führen und einige Elemente von Nahtodeserlebnissen hervorrufen. Auch das soll über eine Beeinflussung der Serotonin-Aktivität geschehen.
1.6
Die euphorisch/psychedelische Wirkung von LSD hängt vom rechten Temporallappen ab. Nach chirurgischer Entfernung des Schläfenlappens können selbst starke Halluzinogene wie LSD keine visuellen Erscheinungen mehr auslösen.
1.6, 3.3 S.44
Mehrere Autoren fanden auch bei einer großen Anzahl von Poltergeist-Fällen bei der den Spuk auslösenden Person eine hohe Inzidenz von epileptischen Störungen, was als Hinweis auf eine epileptoide Basis auch paranormaler psychokinetischer Leistungen zu interpretieren ist.
1.6
Viele Temporallappen-Epilepiker machen Deja-vu-Erfahrungen von etwas, das ihnen völlig vertraut erscheint, obwohl das nicht zutrifft. Diese lassen sich manchmal durch elektrische Stimulation des Temporallappens hervorrufen.
1.6, 3.5.2
Einige haben OBEs, sehen Erscheinungen toter Freunde und Verwandter oder berichten über paranormale Erfahrungen.
3.5.2
Veränderungen der Zeitwahrnehmung können Zeichen einer Schläfenlappenepilepsie sein.
1.6
Die Temporallappenepilepsie führt zu verschiedenen visuellen Phänomenen wie verschwommener Sicht, Hemianopsie (einer Blindheit des halben Gesichtsfeldes), völliger Blindheit sowie in groben Lichtblitzen, verschiedenfarbige Lichtvisionen.
1.6, 3.3 S.246
Das Nahtodeserlebnis unterscheidet sich in einigen Punkten deutlich von einer Schläfenlappenepilepsie, hat aber auch einige Ähnlichkeiten
Nahtodeserlebnisse treten ähnlich plötzlich wie ein epileptischer Anfall auf und haben auch eine anfallsartig kurze Dauer. Das spricht für eine Art epileptoider (=epilepsieähnlich) Entladung, die zu einer sehr schnellen Entwicklung von Bildern oder Halluzinationen führt. Nahtodeserlebnisse sicher keine (temporalen) Epilepsien, da sie sich von diesen in vielem unterscheiden. So sond bei außerkörperlichen Erlebnissen im EEG keine epileptoiden Potentiale vorhanden.
1.6, 1.10
Während epileptischer Anfälle aller Art, besondere aber solcher des Schläfenlappens, fällt die Leistungsfähigkeit beim logischen Denken und beim Erinnern deutlich ab, während das Nahtodeserlebnis zu einer eine enorme Steigerung derselben führt. Tatsächlich zeigen sich während induzierter Außerkörperlichen Erlebnissen in entsprechenden EEG-Ableitungen keine epileptoiden Potentiale.
1.6
Psychose ist die Bezeichnung für verschiedene Formen psychischer Erkrankungen, die einen starken Abbau der Persönlichkeit zur Folge haben. Schläfenlappenepilepsie weist oft psychotische Züge auf. Nahtodeserfahrungen tragen dagegen zur gesunden Weiterentwicklung der Persönlichkeit bei.
1.6
Bei epileptischen Anfällen des Schläfenlappens treten bizarre Körperwahrnehmungsstörungen, Gefühle der Furcht und und automatische Bewegungen auf. Hinzu kommen Schmerzen, Krämpfe, Schwindel und abrupte Gefühlsausbrüchen oder déjà-vus, sowie manchmal auch (gesteigerten) sexuelle Empfindungen. All das kommt in Nahtodeserlebnissen nicht vor. Schließlich sind die optischen Halluzinationen im Rahmen von Schläfenlappenepilepsien im Gegensatz zu den komplexen Bildern von Nahtodeserlebnissen eher bizarr und fragmentarisch, und die typischen Geruchs- und Geschmackshalluzinationen zu Beginn eines solchen Anfalls fehlen im Nahtodeserlebnis völlig.
1.6
Mit ihren gelegentlich Nahtodeserfahrungs-ähnlichen Symptomen liefern Epilepsien Hinweise darauf welche Hirnareale beim Nahtodeserlebnis eine Rolle spielen. Insgesamt ist ein epileptischer Anfall des Schläfenlappens eine ungeordnet auftretende Störung bestimmter Areale des temporolimbischen Systems. Dagegen deutet die Symptomatik der Nahtodeserlebnisse auf eine äußerst genau gesteuerte Aktivation derselben Gehirnbereiche hin.
1.6, 1.10
Bei Lommel wurden mehrere Betroffene erwähnt, die bei der ersten Befragung kurz nach der Wiederbelebung Störungen des Kurzzeitgedächtnisses hatten und sich deshalb an keine Nahtodeserfahrung erinnern konnten, aber bei der zweiten, zwei Jahre späteren Befragung von einem Nahtodeserlebnis berichteten, das sie damals hatten. Das läßt es denkbar erscheinen, daß Erinnerungen an Nahtoderfahrungen nicht während der Erfahrung ins Gehirn gelangen sondern erst danach.
1.1
Nachweise für NTE und Temporallappen
Menschen, die Nahtodeserlebnisse hatten, zeigen mehr Anzeichen von ungewöhnlichen Symptomen, die durch den Schläfenlappen verursacht werden, als die, die in vergleichbaren Situationen keine Nahtodeserfahrung hatten. Es ist jedoch unklar, ob das die Ursache oder die Folge des Nahtodeserlebnisses ist.
1.3
Seifollah Azadi,
Laurie L. Molday, Robert S. Molday: RD3, the protein associated with Leber congenital amaurosis type 12, is required for guanylate cyclase trafficking in photoreceptor cells. In: PNAS, December 7, 2010 107 (49) 21158-21163; (Volltext)
Fritz-Albert Popp:
About the Coherence of Biophotons. "Macroscopic Quantum Coherence", Proceedings of an International Conference on the Boston University, edited by Boston University and MIT, World Scientific 1999. (Volltext 1, 2)
Ein Text von Kersti Nebelsiek, Alte Wilhelmshäuser Str. 5,
34376 Immenhausen - Holzhausen, Tel.: 05673/1615,
https://www.kersti.de/,
Kersti_@gmx.de
Da ich es leider nie schaffe, alle Mails zu beantworten, schon mal
im voraus vielen Dank für all die netten Mails, die ich von
Lesern immer bekomme.
Werbung ist nicht erwünscht und ich bin nicht damit einverstanden, daß diese Adresse für Werbezwecke gespeichert wird.
 Gehirnbereiche, die nötig sind, um Nahtodeserfahrungen zu verarbeiten
Gehirnbereiche, die nötig sind, um Nahtodeserfahrungen zu verarbeiten


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
.png){kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
_2.png){kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}