Was ist Materie? Vom Licht zum komplexen Biomolekül
Inhalt
Übergeordneter Artikel:Dieser Text:
VB218.1 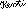 Die Welle-Teilchen-Dualität vom Licht bis hin zum Universum
Die Welle-Teilchen-Dualität vom Licht bis hin zum Universum
VB218.2
Ist das Licht eine Welle, ein Teilchen oder was ganz anderes
VB218.2.1
Einführung
VB218.2.2
Doppelspaltexperiment - Licht als Welle
VB218.2.3
Messungen einzelner Photonen - Licht als Teilchen
VB218.2.4
Funktionsweise der Holographie
VB218.2.5
Nebelkammerexperimente oder die Umwandlung von Licht in Materie
VB218.2.5.1
e = m * c2
VB218.2.5.2
Paarbidungen in der Nebelkammer
VB218.2.5.3
Der Massendefekt im Atomkern
VB218.2.6
Quanten als bewußte Wesen
VB218.3
Atome
VB218.3.1
Atom, das Unteilbare - Atome als Kugeln
VB218.3.2
Bohrsches Atommodell: Atome sind wie eine Sonnensystem! - Kernteilchen und Elektronenbahnen
VB218.3.3
Atome als aus stehenden Wellen aufgebaute Strukturen
VB218.3.3.1
Elektronendiffraktion an Kristallgittern: Das Elektron als Welle
VB218.3.3.2
Unkonventionellere Aspekte des Wassers
VB218.4
Moleküle
VB218.4.1
Moleküle als Stäbchenmodell
VB218.4.2
Moleküle als Kalottenmodell oder Raumfüllendes Modell
VB218.4.4
Aromatische Kohlenwasserstoffe: π-Elektronenring
VB218.4.4
Wasser, Wasserstoffbrückenbindungen und Schneekristalle
VB218.4.4.1
Chemischer und atomphysikalischer Aufbau von Wasser und seinen Kristallen
VB218.4.4.2
Wasser, Wasserstoffbrückenbindungen und Schneekristalle
VB218.5
Die Komplexität biologischer Moleküle
VB218.6
Darstellungen von Proteinen
VB218.6.1
Von der Aminosäure zum Proteinkomplex
VB218.6.2
Die Aminosäuren und ihr Zusammenbau zu Polypeptiden, die Primärstruktur
VB218.6.3
Die Sekundärstruktur des Proteins und eine Einführung in die Bändermodelldarstellung der Proteine
VB218.6.3.1
Die Sekundärstruktur des Proteins
VB218.6.3.2
Alpha-Helix
VB218.6.3.3
Beta-Faltblatt
VB218.6.3.4
Beta-Schleife oder β-Schleife
VB218.6.3.5
Beta-Helix oder β-Helix
VB218.6.3.6
Random-Coil-Strukturen
VB218.6.4
Tertiärstruktur und Quartärstruktur von Proteinen
VB218.6.4.1
Einführung: Tertiärstruktur und Quartärstruktur von Proteinen
VB218.6.4.2
Der ATP-sensitive Kaliumkanal - ein Beispiel für einen Proteinkomplex
VB218.6.4.3
VB218.6.5
Proteinkonformation und Konformationsänderungen
VB218.6.5.1
Einführung: Proteinkonformation
VB218.6.5.2
Aktin und Myosin - Konformationsänderungen, um Bewegung zu erzeugen
VB218.6.5.3
Titin (auch Connectin) im Muskel
VB218.6.5.4
Aktin und Myosin als wesentlicher Teil des Zellskelettes
VB218.6.5.5
Aktin und Myosin in den Mikrovilli
VB218.6.6
Proteine im Gesamtzusammenhang der Zelle: Tubulin und Mikrotubuli
VB218.6.6.1
Vom Tubulin zum Mikrotubulus
VB218.6.6.2
Mikrotubuli als Teil des Zellskelettes
VB218.6.6.3
Mikrotubuli als Eisenbahnschienen der Zelle
VB218.6.6.4
Mikrotubuli in Zilien und Flagellen
VB218.6.6.4.1
Genereller Aufbau von Zilien und Flagellen
VB218.6.6.4.2
Primäre Zilien als Sinnesorgane der Zellen
VB218.6.6.4.3
Sekundäre Zilien oder Flimmerhärchen
VB218.6.6.4.4
Die Flagellen der Eukaryoten
VB218.6.6.5
Teilungsspindel in der Zellteilung
VB218.6.6.6
VB218.6.6.7
Mikrotubuli und Bewußtsein
VB218.6.
VB218.6.
VB218.7
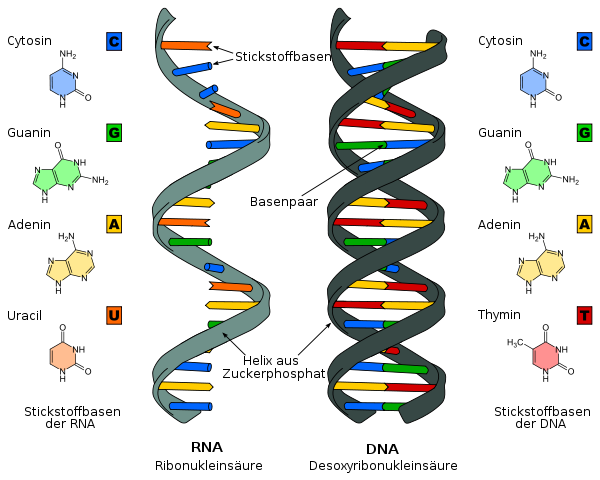
DNA und RNA
VB218.7.1
Nucleoside, die Bausteine von DNA und RNA und der Energiestoffwechsel
VB218.7.1.1
Die Grundstrultur der Nukleobasen, Nukleoside und Nucleotide
VB218.7.1.2
Guanin, Gunanosin und seine Phosphate GDP und GTP
VB218.7.1.3
Adenin, Adenosin, ADP, ATP
VB218.7.1.4
VB218.7.1.5
VB218.7.2
Darstellungen der DNA
VB218.7.3
Verdichtungsgrade der DNA
VB218.7.3.1
Histone und Nukleosomen
VB218.7.3.2
Solenoidstruktur der DNA
VB218.7.3.3
Chromosomenterritorien im Zellkern
VB218.7.3.4
Die Flagellen der Eukaryoten
VB218.
Quellen
10, 20, 30, 40., 50., 60.,70., 80., 90.,110., 120,
1. Die Welle-Teilchen-Dualität vom Licht bis hin zum Universum
Ehe ich davon krank wurde, habe ich einige Jahre Physik und Biologie auf Lehramt studiert. Da ich schon bei Beginn des Studiums älter war als üblich, führte das zu einigen erhellenden Erfahrungen.VA152.
Wie das abgebrochene Studium mein Weltbild verändert hat: Wissenschaft ist ganz anders!Ein Punkt, der mich irritiert hat, war daß ich in der Physik über die Struktur der Materie gelernt hatte, daß letztlich alles Schwingung ist, daß aber in der Biologie gedacht wurde, als wären die Biomoleküle aus Stäbchen und Kugeln zusammengesteckte Molekülmodelle. Während diese Stäbchenmodelle der Moleküle in vieler Hinsicht nützlich sind, sollte man jedoch nicht aus dem Blick verlieren, daß sie eben nur Modellvorstellungen sind, die an manchen Stellen so sehr vereinfachen, daß es schon wieder falsch ist. Außerdem sollte man sich nach Möglichkeit bewußt machen, an welchen Stellen diese dem Modell inhärenten Vereinfachungen falsch sind.
Diese Seite solle eine Hilfesstellung dazu sein, diese Verfälschungen bewußt zu machen.
Wir brauchen Vereinfachungen, um die wesentlichen Aspekte der Realität in unserem Kopf überhaupt unterkriegen zu können, wo diese Vereinfachungen aber Fehler produzieren, sollten wir uns das bewußt machen.
2. Ist das Licht eine Welle, ein Teilchen oder was ganz anderes
2.1 Einführung
Die Frage die den Anstoß zu der Entwicklung der Quantenphysik lieferte war die Welle-Teilchen-Dualität des Lichts. Das Problem daß es sich experimentell nicht entscheiden ließ, ob Licht nun eine Welle ist, oder ein Teilchen.
2.2 Doppelspaltexperiment - Licht als Welle

|
|
Bildquelle: 8.
|

|
|
Bildquelle: 9.
|

|
|
Bildquelle: 10. Daß Wasserwellen solche Interferenzmustererzeugen wird in dem Foto sichtbar, ist aber nicht ganz so deutlich wie bei der Zeichnung. |
VB249.3
Ein Wabenmuster aus Poren im Kondom?VB201.2.3
Chladnische Klangfiguren und was man daraus über Musikinstrumente lernen kann2.3 Messungen einzelner Photonen - Licht als Teilchen
2.4 Funktionsweise der Holographie
|
|
Bildquelle: 1.
|
Es gibt eine Reiher alternativer anordnungen, die ebenfalls holographische Bilder liefern, diese sollen hier aber nicht weiter interessieren, weil diese Variante zum Verständnis des Folgenden ausreicht.
Während ein Photo, das man in der Mitte durchschneidet, in der übriggebliebenen Hälfte genauso detailreich bleibt wie vorher, aber nur noch die Hälfte des Bildes da ist, bleibt bei einem Hologramm, dessen Informationsträger man durchschneidet, das gesamte Bild erhalten, aber es wird deutlich verschwommener, verliert also an Details.
Das Prinzip der Holographie wird nicht nur wie in der obigen Erklärung auf Licht angewandt, sondern es gibt beispielsweise auch eine akustische Holographie, die mit Schallwellen arbeitet und räumliche Klangmuster erzeugt.3.
Die Holographie wird beispielsweise als analogie verwendet, um zu erklären, wie und warum daß ganze Universum als eine Art Hologramm zu verstehen wäre.3.
VB212.3
Quantenverschränkung und die Welt als Hologramm
2.5 Nebelkammerexperimente oder die Umwandlung von Licht in Materie
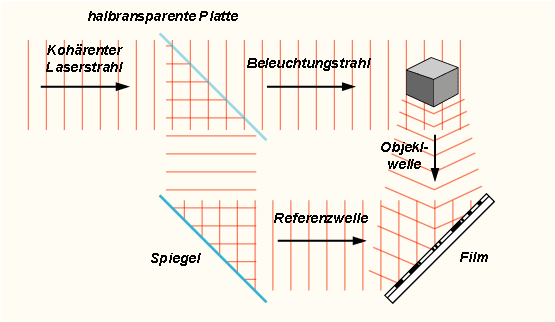
Inhalt2.5.1 e = m * c2
Nach Einstein läßt sich nach folgender Formel wenig Masse in große Mengen an Energie umrechnen.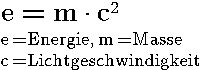
1kg*(3*108m/s)2 = 9*1016(kg m2)/(s2)=9*1016 Ein Kilogramm Masse würde damit 9*1016 Joule entsprechen.
Solarkonstante: 1,39kW/(m2) (Im Vakuum im selben Abstand wie die Erde zur Sonne)
9*1016Ws*3*108m/s ------------------ = 2*1022m3 1,39*103W/(m2)
Wenn Licht in etwa so dicht vorliegt, wie es in der Erdumlaufbahn ist, würde die Menge Photonen, die einem Kilogramm entspricht also einen Würfel mit einer Kantenlänge von etwa 3*104km füllen. Er wäre größer als die Erde.
2.5.2 Paarbidungen in der Nebelkammer
Daß man die Energie des Lichtes tatsächlich in Materie umwandeln kann, ist seit 1933 bekannt. Damals wiesen Frédéric Joliot und Irène Curie das erste mal eine Paarbildung nach, bei der ein Gamma-Teilchen, also ein Photon in ein Paar aus einem Elektron und einem Positron (positiv geladenes Elektron, das Antiteilchen zum Elektron) umgewandelt wird28.. Das Elektron ist mit seiner Ruhemasse von 9*10-31kg29.1 ein sehr leichtes Teilchen, daher ist vergleichsweise wenig Energie nötig, um es zu erzeugen. Es ist so leicht, daß man sein Gewicht in allen Zusammenhängen in denen man die Masse von Neutronen (16726*10-31kg)29.1 und Protonen (16749*10-31kg)29.1 mit 1 u (atomare Masseeinheit, 16606*10-31kg30.) annimmt auf 0 abrundet. Inzwischen wurden auch Antiprotonen und Antineutronen nachgewiesen und es ist klar, daß es zu jedem Elementarteilchen ein Antiteilchen gibt, die, wenn sie zusammentreffen, in ein oder mehrere Photonen zerstrahlen können31..
Frédéric Joliot und Irène Curie das erste mal eine Paarbildung nach, bei der ein Gamma-Teilchen, also ein Photon in ein Paar aus einem Elektron und einem Positron (positiv geladenes Elektron, das Antiteilchen zum Elektron) umgewandelt wird28.. Das Elektron ist mit seiner Ruhemasse von 9*10-31kg29.1 ein sehr leichtes Teilchen, daher ist vergleichsweise wenig Energie nötig, um es zu erzeugen. Es ist so leicht, daß man sein Gewicht in allen Zusammenhängen in denen man die Masse von Neutronen (16726*10-31kg)29.1 und Protonen (16749*10-31kg)29.1 mit 1 u (atomare Masseeinheit, 16606*10-31kg30.) annimmt auf 0 abrundet. Inzwischen wurden auch Antiprotonen und Antineutronen nachgewiesen und es ist klar, daß es zu jedem Elementarteilchen ein Antiteilchen gibt, die, wenn sie zusammentreffen, in ein oder mehrere Photonen zerstrahlen können31..
|
| Bild aus einer Nebelkammer am Deutschen Elektronen Synchroton (DESY), auf dem eine Elektron-Positron Paarbildung farbig markiert ist. Die grün gemalte Bahn des Photons ist im Original nicht sichtbar, sondern wurde aus den Bahnen von Elektron und Positron rekonstruiert 32. |
VB139.
Antiwelten
2.5.3 Der Massendefekt im Atomkern
Daß die atomare Masseeinheit u mit 16606*10-31kg kleiner ist als die Massen von Neutronen (16726*10-31kg)29.1 und Protonen (16749*10-31kg)29.1 ist auf den Massedefekt zurückzuführen. Die atomare Masseeinheit u ist nämlich von dem Kohlenstoffatom mit sechs Protonen und sechs Neutronen abgeleitet und hat ein Zwölftel dieser Masse. Der Unterschied zwischen der Masse der einzelnen Teilchen, aus denen sich das Atom zusammensetzt und dem Atomgewicht ist die Bindungsenergie die frei wird, wenn die einzelnen Teilchen sich zu einem Atom zusammensetzen29. S.821. Je größer diese freiwerdende Energie ist, desto stabiler ist ein Atom, da eben diese Energiemenge nötig ist, um das Atom zu spalten. Auch in diesem Zusammenhang tritt also eine Umwandlung von Energie in Masse und umgekehrt auf. Dies wird bei der Kernspaltung und der Kernfusion ausgenutzt.
Ausgenutzt habe ich das um den Sinn des Begriffes "feinstofflich" zu erklären.
VA309.
Ist Feinstoffliches materiell?
2.6 Quanten als bewußte Wesen
In "The Lenses of Perception Interpretation of Quantum Mechanics" vergleicht Doug Marman das Verhalten von quanten mit den Entscheidungen empfindungsfähiger Wesen und kommt zu dem Schluß, daß Quanten so reagieren wie empfindungsfähige Wesen, indem sie Beziehungen eingehen (Quantenverschränkung) und diesen Beziehungen entsprechend handeln133.. Die über hundertseitige Erklärung bekommen ich hier nicht in einem kurzem Absatz zusammengefaßt, doch finde ich das vor allem auch deshalb interessant, da meine feinstofflichen Anteile der Ansicht sind, daß die kleinsten Teilchen aus denen Materie entstanden ist, winzige abgespaltene Anteile weitaus größerer Wesen sind. Sie haben also aus einer völlig anderen Perspektive heraus auch behauptet, daß Quanten bewußte Wesenheiten sind.VB99.2.1.5
Zusammengeballter Staub: Die Materie entsteht
3. Atome
3.1 Atom, das Unteilbare - Atome als Kugeln
Das Wort Atom ist von altgriechisch ἄτομος átomos‚ unteilbar abgeleitet. Dies geht auf die Vorstellung zurück, Materie wäre aus unteilbaren Elementarteilchen, den Atomen, aufgebaut, die man sich als kleine Kugeln vorstellem könne.Davon abgeleitet ist die heute noch verwendete Darstellung von Molekülen als Kugeln die mit Stäbchen, die für Atombindungen stehen, verbunden sind und die Darstellungen von Kristallen als regelmäßig angeordnete Strukturen.
3.2 Bohrsches Atommodell: Atome sind wie eine Sonnensystem! - Kernteilchen und Elektronenbahnen
Als genug Atome und ihre Eigenarten bekannt waren stellte sich heraus, daß Atome, wenn man sie nach ihrem Atomgewicht sortiert periodisch wiederkehrende Eigenarten haben. Dies führte zu der Erstellung des Periodensystems der Elemente.
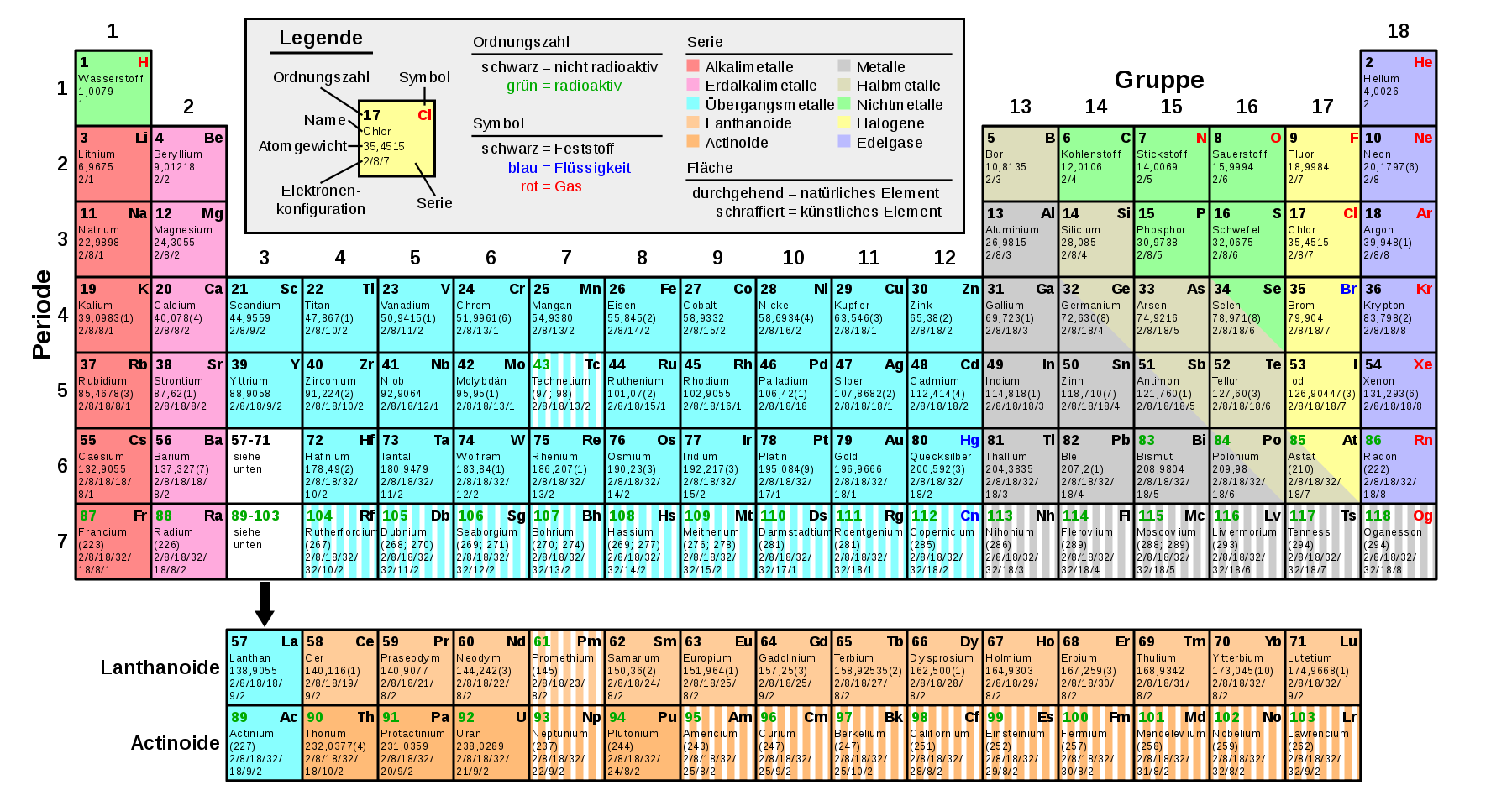
|
|
Bildquelle: 7. |
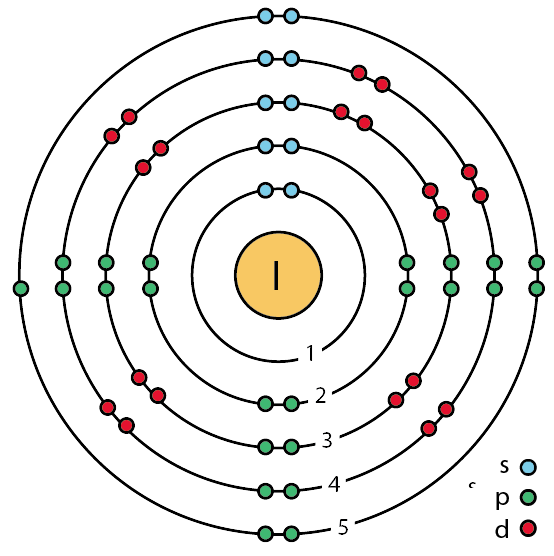
|
|
Bildquelle: 11. Bohrsches Atommodell von Jod (Ordnungszahl 53) |

|
|
Bildquelle: 13. Absorptionsspektrum und Emissionsspektrum von Wasserstoff: Dieselben Wellenlinien die Wasserstoff absorbiert strahlte es im angeregten Zustand auch ab, nämlich 410nm, 434nm, 486nm, 656nm |
|
|
Bildquelle: 12. Im Rahmen des Bohrschen Atommodells, läßt sich die Absorption dadurch erklären, daß ein Elektron auf eine höhere Elektronenbahn angehoben und dafür die Energie des Lichtquants verbraucht wird. Derselbe Energiebetrag wird wieder frei, wenn das Elektron wieder auf eine tiefere Elektronenbahn abfällt. |
|
|
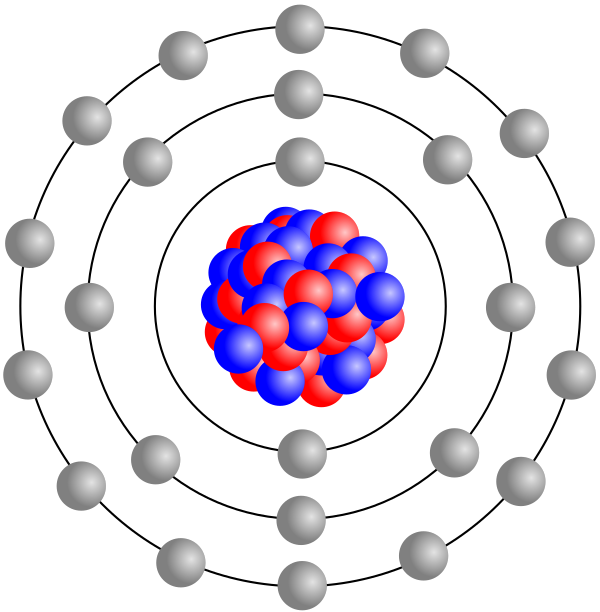
Bildquelle: 17. Eisenatom mit Darstellung der Protonen, Neutronen und Elektronen als kleine Kügelchen. |
3.3 Atome als aus stehenden Wellen aufgebaute Strukturen:
3.3.1 Elektronendiffraktion an Kristallgittern: Das Elektron als Welle
Louis de Broglie (1892-1987) war der Ansicht, daß man die Welle-Teilchen-Dualität des Lichtes ernst nehmen und daher dem Licht eine Ruhemasse zuordnen könne, die seiner aus Einsteins Formel abgeleiteten Masse entspricht.
Umgekehrt leitete er davon aber auch eine Wellenlänge für materielle Teilchen ab, schreib also allen Teilchen Welleneigenschaften zu.37. S.20
Clinton Joseph Davisson und Lester Halbert Germer34. sowie George Paget Thomson und Alexander Reid35., 36. wiesen zuerst nach, daß Elektronendiffraktion - Interferenzen durch Beugung am Atomgitter - auftreten, wenn man einen Elektronenstrahl durch dünne Folien aus kristallinem Material hindurchschickt. Damit war nachgewiesen, daß Elektronen Welleneigenschaften besitzen, Broglies Formel also tatsächlich für sie gilt.
3.3.2 Elektronenorbitale - Wellenformeln erklären die Struktur der Materie
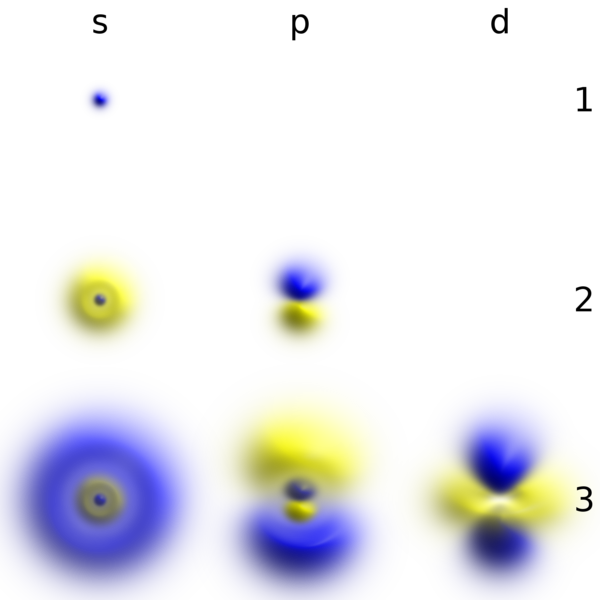
Die Elektronenorbitale werden berechnet, indem man die Wellenfunktion der Schrödingergleichung auf die Elektronen eines Atoms anwendet. Das Elektron wird in diesem Falls also nicht als Teilchen sondern als Welle betrachtet. Interpretiert man das dann wiederum auf der Teilchenebene, beschreibt das Elektronenobrital die Aufenthaltswahrscheinlichkeit des Elektrons in dieser Elektronenbahn.
|
|
Bildquelle: 14. Für das einzige Elektron des Wasserstoffatoms wurden hier die Lösungen der Schrödingergleichung für verschiedene Elektronenorbitale berechnet, die die Nummern 1s, 2s, 2p, 3s, 3p and 3d tragen. Normalerweise befindet sich das Elektron im 1s Orbital, das die Form einer kleinen Kugel hat, wie oben links dargestellt. Die anderen Orbitale werden nur in angeregten Zuständen genutzt. Aus den Berechnungen der mögliche Elektronenbahnem im Wasserstoffatom läßt sich auch die Frequenz der Linien im Absorptionsspektrum und Emissionsspektrum von Wasserstoff, das oben dargestellt ist berechnen. |

|
|
Bildquelle: 15.
Stehende Wellen auf eindimensionalen schwingenden Medien, den Guitarrensaiten: |

|
|
Bildquelle: 16.
Stehende Wellen auf zweidimensionalen schwingenden Medien |
4. Moleküle
4.1 Moleküle als Stäbchenmodell
Obwohl wir schon lange wissen, daß Atome keine Kugeln sind, wird die räumliche Struktur von Molekülen immer noch gerne so dargestellt, als wären Atome tatsächlich Kugeln, die durch Atombindungen verbunden sind, die wie Stäbchen dargestellt werden.Daß man das so macht, liegt daran, daß weder das Bohrsche Atommodell noch das Orbitalmodell besonders hilfreich sind, wenn man komplexe Moleküle darstellen will, weil wir, wenn wir so viele Details einzeichnen, schlicht den Überblick verlieren.

|
|
Bildquelle: 5. Kristallstruktur von Bergkristall als Stäbchenmodell, es handelt sich hierbei um Siliziumdioxid (SiO2), Sauerstoff ist gelb, Silizium rot dargestellt. Es ist erkennbar wie die Form des Kristalls entsteht. |

|
|
Bildquelle: 6. Bergkristall |
4.2 Moleküle als Kalottenmodell oder Raumfüllendes Modell
Die Kalottenmodelle haben folgende anderen Namen: Corey-Pauling-Koltun-Modelle, C-P-K-Modelle, CPK-Modelle, englisch: space-filling atomic models27..
Robert B. Corey und Linus Pauling entwickelten 1952 das Kalottenmodell, um anschauliche räumliche Molekülmodelle zum Zusammenstecken bauen zu können25., nachdem er die Steckverbindung dazu optimiert hatte33., kündigte Walter L. Koltun 1965 an, daß die Modellbausteine bald käuflich zu erwerben sein werden.27.
Die Form der Einzelteile beruht auf festgestellten Atomabständen in Kristallgittern, die sich beispielsweise aus der Untersuchung durch Diffraktion an Kristallgittern in Rönthgenuntersuchungen nachweisen ließen38., 39.. Diese Abstände werden zur Berechnung der van der Waals radii und Bindungslängen herangezogen. Die einzelenen Atommodelle sind so konzipiert, daß ser gesamte Raum zwischen zwei Atomkernen ausgefüllt ist, also das Atom samt Atomhülle als Kugel dargestellt wird, die da abgeschnitten ist, wo sich ein anderes Atom daran binden könnte. 24., 25., 27.

|
|
Bildquelle: 134. Raumfüllendes Modell oder Kalottenmodell von LSD |
4.3 Aromatische Kohlenwasserstoffe: π-Elektronenring
Nach dem giechischen Buchstaben π (Pi) ist eine Struktur benannt, die in aromatischen Kohlenwasserstoffen auftaucht, der π-Elektronenring.
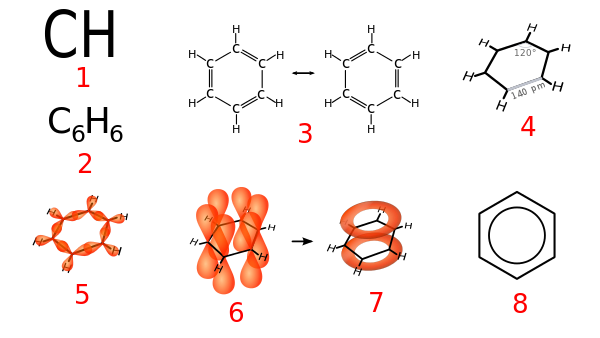
|
|
Bildquelle: 4. Verschiedene Darstellungen des Benzens:
|
4.4 Wasser, Wasserstoffbrückenbindungen und Schneekristalle
4.4.1 Chemischer und atomphysikalischer Aufbau von Wasser und seinen Kristallen
Wasser hat die Summenformel H2O. Wasser ist im Vergleich zu nahezu allen anderen einfachen Verbindungen über einen ausgesprochen großen Temperaturbereich hinweg flüssig. Die Ursache dafür findet sich in der Struktur des Moleküls.
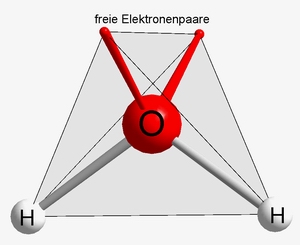
|
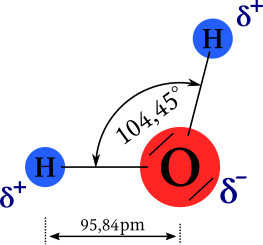
| |
|
Bildquelle: 68. räumlicher Aufbau eines H2O-Moleküls. Die freien Elektronenpaare des Sauerstoffatoms bilden zusammen mit den H-Atomen ein Tetraeder. Hieraus ergibt sich, warum das Wassermolekül nicht linear aufgebaut ist sondern etwas gebogen. |
Bildquelle: 69. Der Bindungswinkel zwichen den beiden Bindungen der Wasserstoffmoleküle beträgt, 104,45°, die Bindungslänge 95,84pm. Da das Sauerstoffatom die Elektronen etwas stärker anzieht als die Wasserstoffatome ist das Wassermolekül auf der Sauerstoffseite schwach negativ gelasten auf der Seite der Wasserstoffatome ist es schwach positiv geladen. |

|
|
Bildquelle: 70. Diese Ladung, die deutlich schwächer ist als die negative Ladung eines einzelnen Elektrons, führt dazu daß sich im flüssigen Wasser sogenannte Wasserstoffbrückenbindungen (gepünktelte Linien) ausbilden, die schwächer sind als normale Atombindungen, aber doch bewirken, daß das Wasser früher vom gasförmigen in den flüssigen Zustand übergeht als die meisten Gase. |
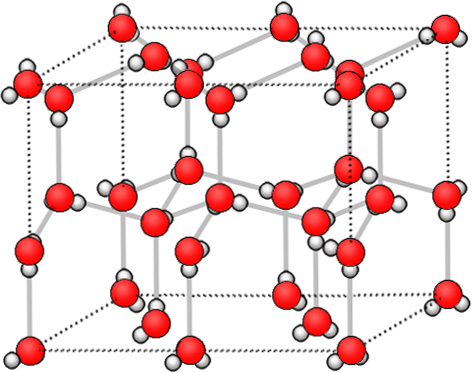
|
|
Bildquelle: 71. Die Wasserstoffbrückenbindungen (grau) sind auch für die Kristallstruktur von Eis verantwortlich. |
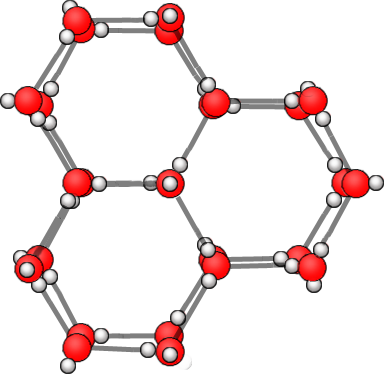
|
|
Bildquelle: 72. Wenn man die Kristallstruktur etwas dreht, wird deutlich, woher die sechseckige Grundstruktur der Schneekristalle kommt.
|

|
|
Bildquelle: 73. Photomontage aus Schneeflockenfotos von Wilson Bentley (1865-1931). |
4.4.2 Unkonventionellere Aspekte des Wassers
2002 fand ich in einer Buchhandlung Bücher von Masaru Emoto65., 66., 67., sah mir die Bilder an, fand "Oh sind die schön!" und mußte sie deshalb alle haben. Die Bücher enthielten diverse Bilder von Wasserkristallen und handelten davon, daß die Form der Kristalle, die gefrieren entstehen von diversen Faktoren abhängen, unter anderem:
- Der Herkunft des Wassers - ob es beispielsweise aus Heilquellen, Gebieten mit erheblicher Wasserverschmutzung, sauberen Gegenden stammt
- Nachfolgender Behandlung mit Musik, durch Destillation etc.
- körperliche und psychische Verfassung der Menschen, die mit dem Wasser umgehen
Manchmal habe ich den Eindruck, es läge daran, daß das Thema "Iiii!" wäre, manchmal daß es zu kompliziert ist - schließlich bedeuten so viele Einflußfaktoren auch, daß man schwer trennen kann, was genau woher kommt. Jedenfalls kam diese Forschung von Emoto nicht wirklich in der Mainstream-Forschung an.
Daß aber Informationen in Materie auf eine Weise gespeichert werden, die anhand seiner Chemie nicht zu verstehen ist, ist auch aus einer anderen Richtung bekannt, nämlich aus der Homöopathie.
VA193.
Homöopathie: Kann eine homöopathische Behandlung helfen, obwohl homöopathische Potenzen keinen Wirkstoff enthalten?

|
|
Bildquelle: 74.
|
5. Die Komplexität biologischer Moleküle
Ende 2019 wunderte ich mich, warum ich plötzlich so fasziniert von den Zellwandstrukturen der Bakterien war.VB198.
Bakterien und andere Prokaryoten sind bereits hochkomplexe WesenDarauf meldeten sich Anteile von mir, die früher einmal Bakterien gewesen waren und sagten
"Wir haben doch damals so tolle Sachen erfunden. Das wollen wir aufschreiben."
Mit damals meinten sie, wie sie mir zeigten, das Präkambrium.
T1. 4 500 - 600 Mio Präkambrium
Während die grundlegenden Erfindungen wohl von damals stammten, habe ich in meinem Bakterienartikel natürlich moderne Bakterien beschrieben, wie es sie heute gibt.
Meine jetzige Beschäftigung mit dem Thema paßt in dieses Bild:
Bei der Beschäftigung mit diesen Biomolekülen hatte ich den Eindruck, es mit faszinierenden kleinen Maschinchen zu tun zu haben, die wie winzige technische Geräte funktionieren und in denen jedes einzelne Atom eine bestimmte Funktion hat. Das könnte einem in Bezug auf das Thema Nanobots zu denken geben...
VB216.
Gibt es Nanobots, mit denen die Geheimgesellschaften die Menschheit manipulieren?
6. Darstellungen von Proteinen
6.1 Von der Aminosäure zum Proteinkomplex
Proteine, umgangssprachlich Eiweiße sind biologische Makromoleküle, die sich aus langen Ketten von Aminosäuren zusammensetzen, die wiederum zu noch komplexeren Gebilden zusammengebaut sind. Die Struktur von Eiweißen hat mehrere aufeinander aufbauende Komplexitätsstufen.- Primärstruktur: Reihenfolge der Aminosäuren
- Sekundärstruktur: Zusammensetzung des Proteins aus besonders häufig auftretenden Motiven für die räumliche Anordnung der Aminosäuren
- Tertiärstruktur: Anordnung der Sekundärstrukturen zu komplexen dreidimensionalen Formen
- Quartärstruktur: Zusammensetzung eines größeren Komplexes aus mehreren in ihrer Tertiärstruktur gefalteten Proteinen
6.2 Die Aminosäuren und ihr Zusammenbau zu Polypeptiden, die Primärstruktur
Eine Aminosäure Ist ein Molekül, das folgende grundsätzliche Struktur hat.
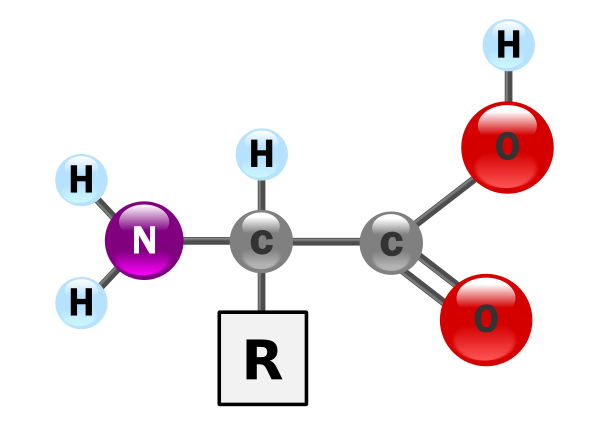
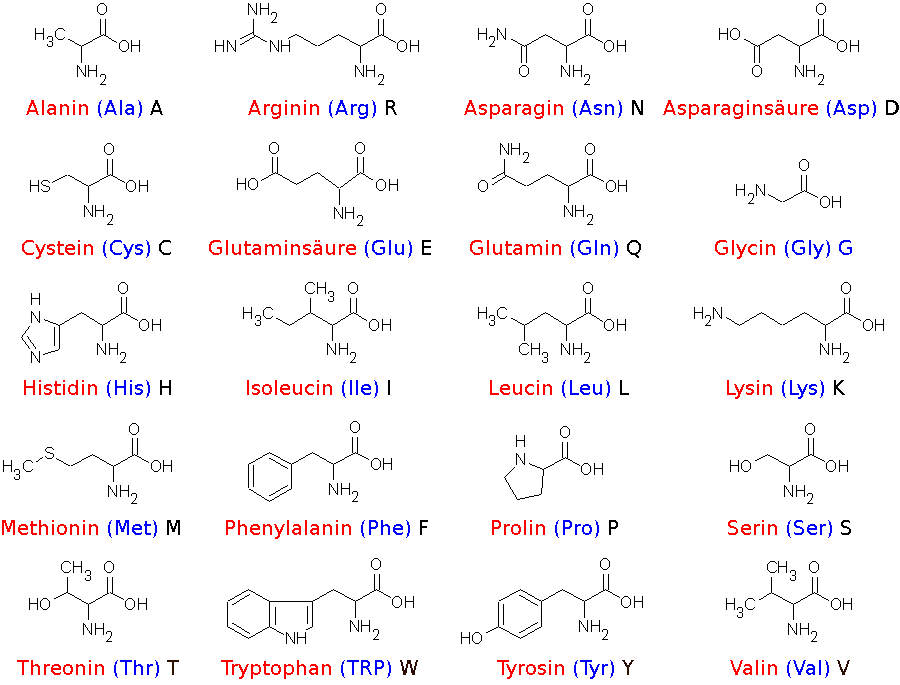
|
|
Bildquelle: 44. Strukturformeln der 20 Aminosäuren ohne Angabe der räumlichen Anordnung der Atome im Molekül.
Blau: Drei-Buchstaben-Kürzel der Aminosäure Schwarz: Einbuchstabencode
|
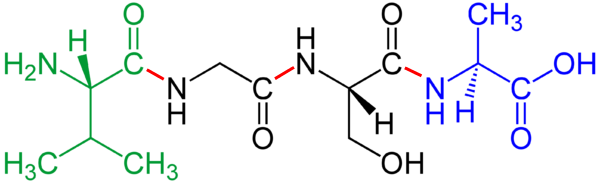
|
|
Bildquelle: 46. Kette aus 4 Aminosäuren, die Bindung zwischen zwei Aminosäuren wurde von mir rot markiert und ist die Stelle, wo das Wasser - beziehungesweise eine OH-Gruppe auf der einen und ein Wasserstoff (H) auf der anderen Seite - abgespalten und durch eine Atombindung ersetzt wurde. |
6.3 Die Sekundärstruktur des Proteins und eine Einführung in die Bändermodelldarstellung der Proteine
6.3.1 Die Sekundärstruktur des Proteins
Wenn man bedenkt, daß 100 bis 300 Aminosäuren eine Polypeptidkette ergibt, ist offensichtlich, daß es zu unübersichtlich wäre, jedes Atom einzeln einzuzeichnen. Umd Peptide übersichtlicher darzustellen wurde daher das Bändermodell erfunden. Einige häufige Strukturelemente haben im Bändermodell besondere Darstellungsweisen. Man unterscheidet dabei zwischen folgenden Strukturtypen: α-Helix, β-Faltblatt, β-Schleife, β-Helix und ungeordnete, so genannte Random-Coil-Strukturen. Insgesamt besteht die Aufgabe des Bändermodells darin, einen räumlichen Eindruck der Form eines Moleküls zu erzeugen, der die innere Struktur ebenfalls übersichtlich wiedergibt.
6.3.2 Alpha-Helix oder α-Helix
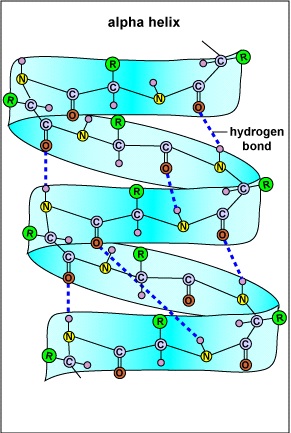
|
|
Bildquelle: 47.
Alpha Helix |
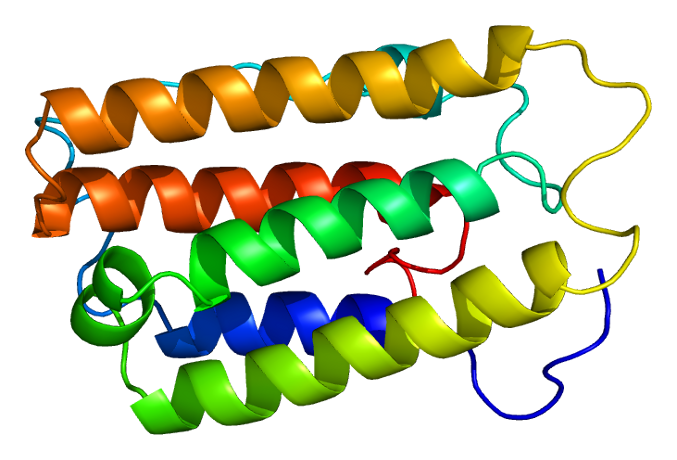
|
|
Bildquelle: 48. Bändermodell von Interferon alpha-2, einem Protein mit 5 Alpha-Helices. Es spielt bei der Abwehr von Viren und Bakterien eine Rolle. Um trotz der verdeckten Stellen deutlich zu machen, in welcher Reihenfolge die einzelnen Teile der Polypeptidkette zusammenhängen, wurde das Bändermodell des Polypeptids in der Reihenfolge der Regenbogenfarben angefärbt. |
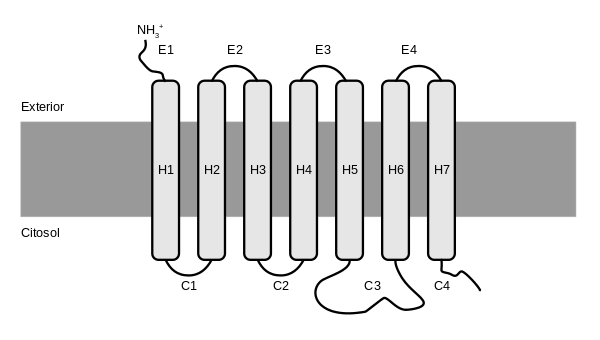
|
|
Bildquelle: 119. Struktur eines G-Protein-gekoppelten Rezeptors - Transmembrandomänen in einer Reihe in die Membran gezeichnet |
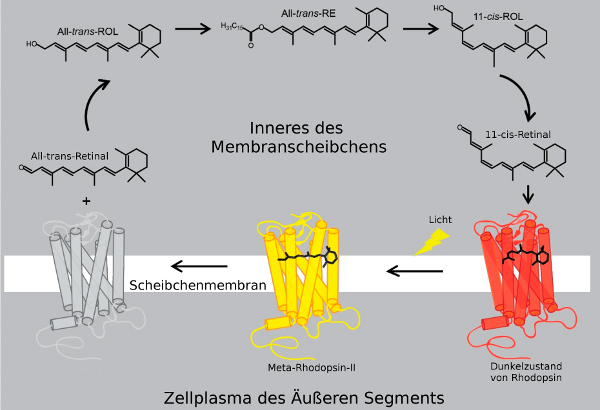
|
|
Bildquelle: 132.
Um die Lichtaktivierung des Rhodopsins und das Recycling von Retinal darzustellen, wurde hier der G-Protein gekoppelter Rezeptor Rhodopsin in einer räumlichen Zylinderdarstellung abgebildet. Während die Form der einzelnen α-Helices nur sehr schematisch dargestellt ist, ist gezeigt, daß sich das Retinol bei der Lichtaktivierung streckt und die einzelenen Helices dadurch auseinanderschiebt. Die genauere Erklärung findet sich hier: |
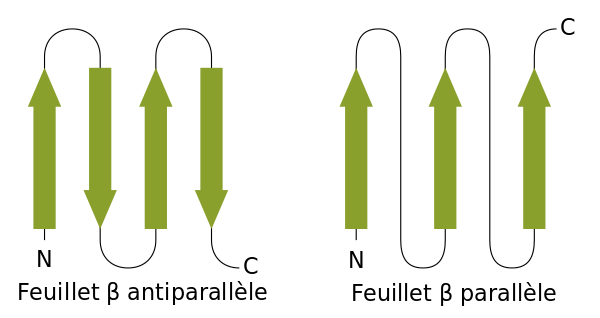
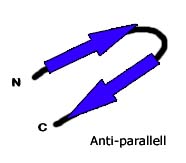
6.3.3 Beta-Faltblatt oder β-Faltblatt
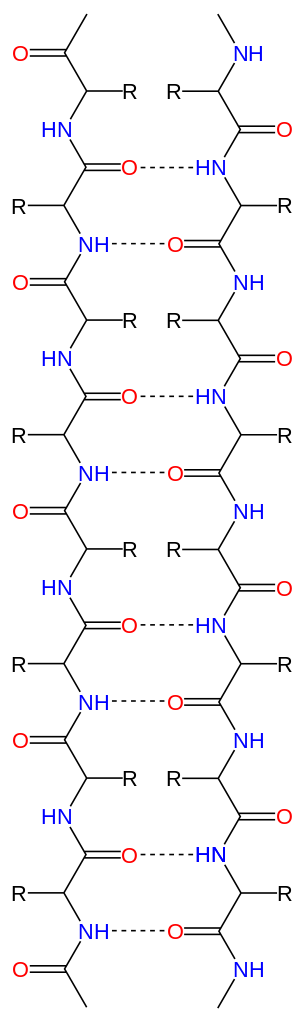
|
|
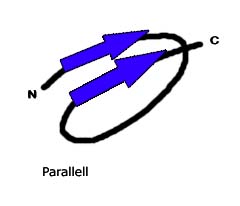
Bildquelle: 49. Antiparalleles Beta-Faltblatt: Durch Wasserstoffbrückenbindungen (gestrichelte Linien) zwischen dem Sauerstoff (rot) und dem an den Stickstuff gebundenen Wasserstoff (blau) wird die Struktur des Beta-Faltblattes zusammengehalten Im Bändermodell wird das antiparallele Beta-Faltblatt durch zwei entgegengesetzt gerichtete Pfeile dargestellt |
|
|
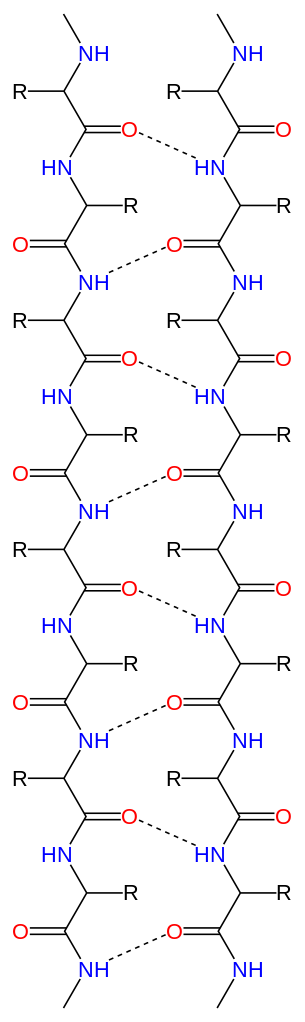
|
|
Bildquelle: 50. Paralleles Beta-Faltblatt: Durch Wasserstoffbrückenbindungen (gestrichelte Linien) zwischen dem Sauerstoff (rot) und dem an den Stickstuff gebundenen Wasserstoff (blau) wird die Struktur des Beta-Faltblattes zusammengehalten Im Bändermodell wird das parallele Beta-Faltblatt durch zwei gleich gerichtete Pfeile dargestellt |
|
|
|
|
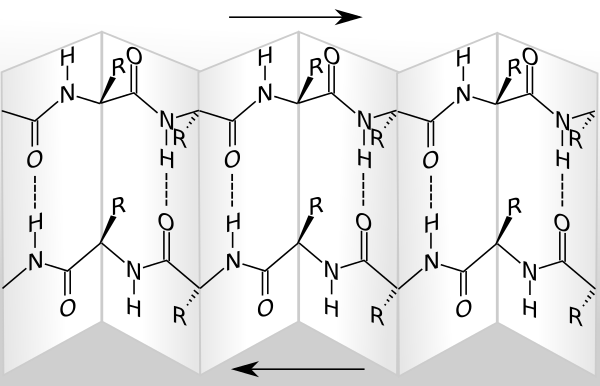
Bildquelle: 51. Antiparalleles Beta-Faltblatt: Als Faltblatt wird das Beta-Faltblatt bezeichnet, weil es nicht flach ist wie die beiden oberen Zeichnungen nahelegen würden, sondern so gefaltet ist, daß der eine Rest nach oben, der nächste aber nach unten raussteht. |
|
|
Bildquelle: 54. Sowohl das parallele als auch das antiparallele Beta-Faltblatt kann zu größeren Flächen zusammengesetzt werden, die dann durch entsprechend mehr Pfeile dargestellt werden |
|
|
Bildquelle: 55.
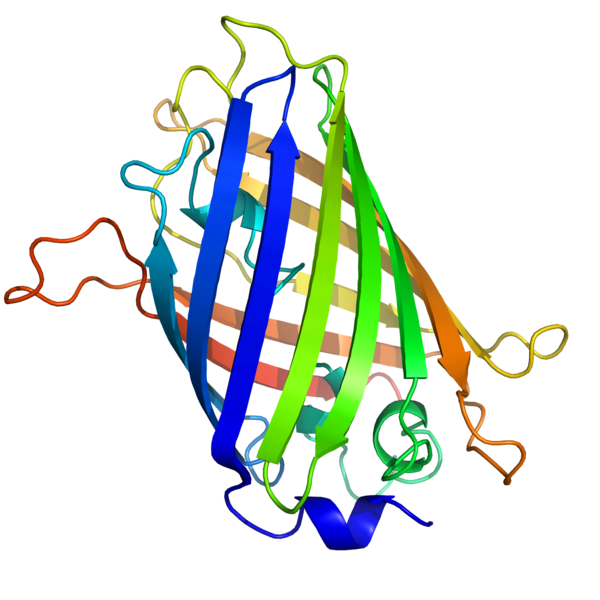
Ein Beispiel für ein Protein mit mehreren antiparallelen Beta-Faltblättern ist das grün fluoreszierende Protein (Green Fluorescent Protein, kurz: GFP) der Qualle Aequorea victoria, das beispielsweise in der Mikroskopie verwendet wird, um mit Antikörperfärbungen mikroskopische Strukturen gezielt zu markieren und sichtbar zu machen, wo sie zu finden sind.56. |
Beim AcrAB-TolC-Effluxpumpensystem von Escherichia coli enthält das äußere Membranprotein TolC ein β-Fass.
VB198.3.3.2
Das AcrAB-TolC-Effluxpumpensystem von Escherichia coli
6.3.4 Beta-Schleife oder β-Schleife
Anderen Bezeichnungen: β-Kehre, englisch β-Turn, Haarnadelschleife oder Haarnadelkehre
|
|
Bildquelle: 57. β-Schleife |
|
|
Bildquelle: 52. Bei der hier dargestellten Kurve im antiparallelen Beta-Faltblatt könnte es sich um eine β-Schleife handeln, muß es aber nicht. |
6.3.5 Beta-Helix oder β-Helix
Eine β-Helix besteht aus mehreren parallelen β-Strängen, ähnlich dem parallelen Beta-Faltblatt. Die Polypeptitkette hat insgesamt die Form einer Spirale, von der mindestens eine Seite durch dieses Beta-Faltblatt gebildet wird. Es können aber auch mehrere solche Beta-Faltblatt-Seiten in die Spirale eingebaut sein.

|
|
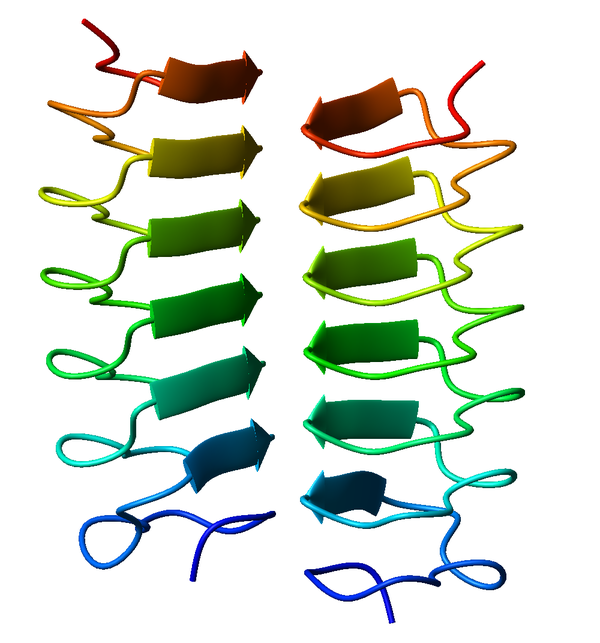
Bildquelle: 58. Bändermodell eines Frostschutzproteins aus (Choristoneura fumiferana). Drei Seitenflächen der Spirale werden durch parallele Beta-Faltblattstrukturen gebildet. Die Moleküle setzen sich auf die Stellen, wo sich Eis zu bilden beginnt und verhindern so, daß die Eiskristalle auf eine nennenswerte Größe anwachsen können.138. |

|
|
Bildquelle: 135. Puppe und Falter von Choristoneura fumiferana auf einer Fichtenart (Picea sp.). Es handelt sich um eine nordamerikanische Schmetterlingsart aus der Familie der Wickler (Tortricidae). |

|
|
Bildquelle: 136. Eier von Choristoneura fumiferana. Die Raupen schlüpfen noch im Herbst und verstecken sich unter der Borke oder an anderen geschützten Stellen auf dem Wirtsbaum. Sie können mit Hilfe des Frostschutzproteins sehr tiefe Temperaturen bis zu minus 30 Grad Celsius überleben138., wenn es aber zu lange kalt ist, verhungern sie. |

|
|
Bildquelle: 137. Ältere Raupe von Choristoneura fumiferana, wie sie im Sommer zu sehen sind. Sie fressen gewöhnlich an Balsam-Tanne (Abies balsamea), der Weiß-Fichte (Picea glauca) und der Amerikanischen Rot-Fichte (Picea rubens) |
|
|
Bildquelle: 59. Bändermodell eines Frostschutzproteins des Mehlkäfers (Tenebrio molitor) in zwei verschiedenen Ansichten. Eine Seitenfläche der Spirale wird durch parallele Beta-Faltblattstrukturen gebildet. |
6.3.6 Random-Coil-Strukturen
Als Random-Coil-Strukturen werden Proteinstrukturen bezeichnet, die sich nicht in die häufigen und bekannten Muster einordnen lassen sondern irgendwie anders sind. Sie werden im Bändermodell meist als dünne Drähte dargestellt.
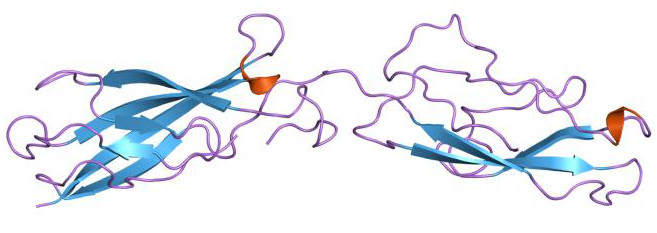
|
|
Bildquelle: 60.
N-Cadherin (neuronales Cadherin) der Maus (Mus musculus). Es stimuliert über Zell-Zell-Kontakte die Migration und Invasion von Zellen und spielt damit eine bedeutende Rolle in der Embryonalentwicklung. |
6.4 Tertiärstruktur und Quartärstruktur von Proteinen
6.4.1 Einführung: Tertiärstruktur und Quartärstruktur von Proteinen
Beispiele für Tertiärstrukturen von Polypeptiden habe ich weiter oben schon gegeben. Das Bändermodell von Interferon alpha-2 zeigt wie fünf 5 Alpha-Helices in der Proteinstuktur eines einzelnen Polypeptits angeordnet sind.VB218.6.3.2
Alpha-HelixDas grün fluoreszierende Protein (Green Fluorescent Protein, kurz: GFP) der Qualle Aequorea victoria habe ich als Beispiel für die Tertiärstruktur eines Polypeptides gewählt, das Beta-Faltblätter enthält.
VB218.6.3.3
Beta-FaltblattBei den beiden Frostschutzproteinen als Beispiele für die Beta-Helix ist die Sekundärstruktur praktisch mit der Tertiärstruktur identisch, weil sie nur aus eben dieser Beta-Helix bestehen.
VB218.6.3.5
Beta-Helix oder β-HelixN-Cadherin dient als Beispiel für die Tertiärstruktur eines Polypeptides, das relativ viele Random-Coil-Strukturen enthält.
VB218.6.3.6
Random-Coil-StrukturenAll diese Beispiele bestehen aus einer einzigen Polypeptidkette. Sie haben daher keine Quartärstruktur.
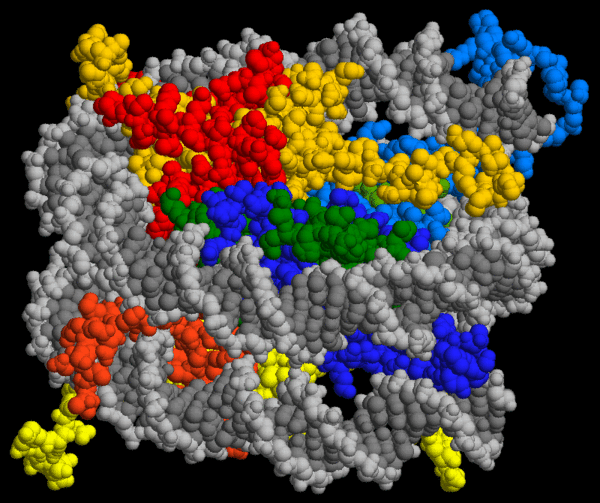
Viele Proteine sind komplex zusammengesetzte Strukturen aus mehreren in ihrer Tertiärstruktur gefalteten Polypeptidketten, sogenannte Proteinkomplexe. Wie sich ein solcher Proteinkomplex aus seinen Einzelteilen zusammenpuzzlet, beschreibt die Quartärstruktur eines Proteins.
6.4.2 Der ATP-sensitive Kaliumkanal - ein Beispiel für einen Proteinkomplex
Die folgenden Bilder zeigen, wie Gregory M. Martin et. Al. mit Kryoelektronenmikroskopie die räumliche Struktur eines Proteinkomplexes ermittelt haben61.. Die Kryoelektronenmikroskopie (Kryo-EM) ist eine Form der Transmissionselektronenmikroskopie (TEM), bei welcher biologische Proben bei kryogenen Temperaturen (≲ −150 °C) untersucht werden.

|
|
Bildquelle: 61.1
ATP-sensitiver Kaliumkanal des Goldhamsters (Mesocricetus auratus)
|

|
|
Bildquelle: 61.2 Der blau dargestellte Innenteil Kir6.2 des ATP-sensitiven Kaliumkanals ist hier noch einmal einzeln zu sehen. Diese Form hat der Kanal, da er verschlossen ist, da viel Glibenclamid (GBC - ein Antidiabetikum, das die ATP-sensitiver Kaliumkanäle verschließt) und Adenosintriphosphat (ATP) vorhanden war.
|
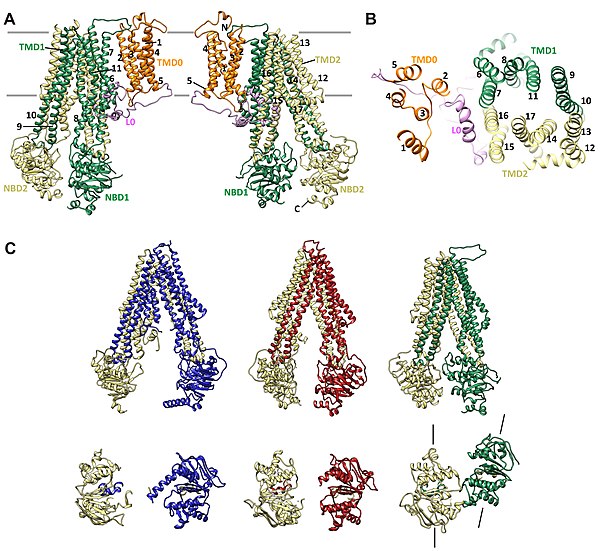
|
|
Bildquelle: 61.3 Der in denselben Farben wie oben dargestellte Außenteil SUR1 des ATP-sensitiven Kaliumkanals ist hier noch einmal einzeln zu sehen. Dieser Kanal gehört zu den ABC-Transportern, öffnet bei Aktivierung mit ATP aber nur den Kaliumkanal, während viele andere ABC-Transporter Biomoleküle von erheblicher Größe aus der Zelle exportieren.
|

|
|
Bildquelle: 61.6 Der etwa 1mm lange Fadenwurm Caenorhabditis elegans unter dem Lichtmikroskop |
VB198.3.3.2
RTX-Proteine und das Typ-I-Sekretion-System (T1SS)Die weiteren Untersuchungen beschäftigen sich mit der Funktion des ATP-sensitiven Kaliumkanals in den B-Zellen der Langerhansschen Inseln des Pankreas. Die Autoren wollen herausfinden, warum das zu den Sulfonylharnstoffen gehörende Glibenclamid den ATP-sensitiven Kaliumkanal verschließt und auf diesem Wege eine Insulinaussschüttung bewirkt.
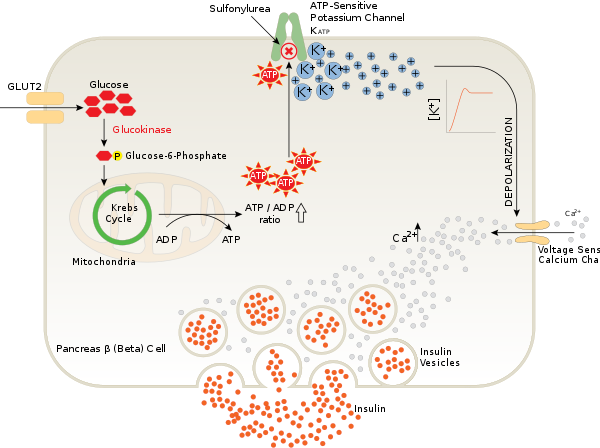
|
|
Bildquelle: 64.
Funktion des ATP-sensitiven Kaliumkanals in den B-Zellen der Langerhansschen Inseln des Pankreas. Durch den Krebs-Zyklus (grüner Kreis) in den Mitochondrien wird diese Energie verwendet, um aus ADP ATP (rote Sterne) herzustellen. Das ATP verschließt den ATP-sensitiven Kaliumkanal (oben, grün), so daß sich K+-Ionen in der Zelle ansammeln und die Zelle depolarisieren. Dies öffnet den spannungsgesteuerten Kalziumkanal (beige, im rechten Rand der Zelle), so daß Ca2+-Ionen (graue Punkte) in die Zelle einströmen, die wiederum bewirken daß die mit Insulin (orangene Punkte) gefüllten Vesikel (graue Kreise) Insulin aus der Zelle ausschütten. |
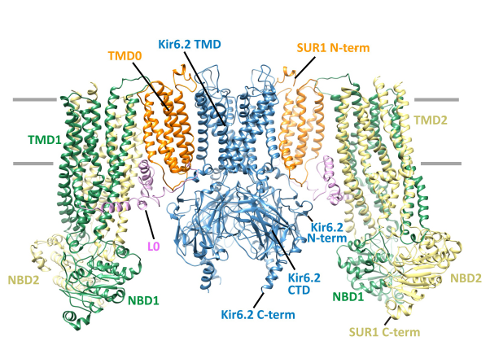
|
|
Bildquelle: Noch einmal das Bändermodell des ATP-sensitiven Kaliumkanals |
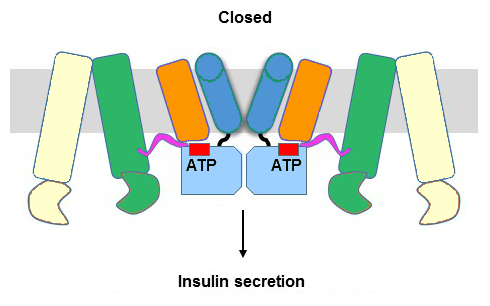
|
|
Bildquelle: 61.5 Eine vereinfachte Darstellung des ATP-sensitiven Kaliumkanals in grob denselben Farben wie oben sähe so aus |
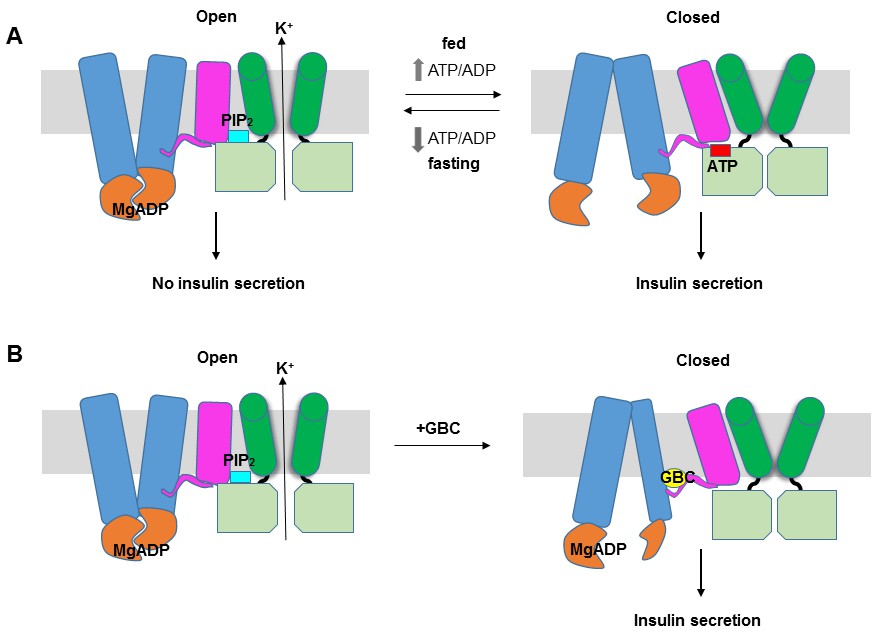
|
|
Bildquelle: 61.4
Funktionsweise des ATP-sensitiven Kaliumkanals |
6.5 Proteinkonformation und Konformationsänderungen
6.5.1 Einführung: Proteinkonformation
Die Gesamtform oder Konformation eines Proteins, wie sie durch Sekundär-, Tertiär- und Quartärstruktur des Proteins entstanden ist, kann sich ändern uns solche Änderungen spielen in verschiedenen Zusammenhängen eine Rolle.
Die Öffnung des ATP-sensitiven Kaliumkanals stellt eine solche Konformationsänderung dar, das heißt der Kanal wird tatsächlich breiter, so daß innen mehr durchpaßt, wenn er geöffnet wird. ein weiteres Beispiel für ein kanalprotein, wo die Proteinkonformation ine Rolle für die Funktion eines proteins spielt behandele ich beim Das AcrAB-TolC-Effluxpumpensystem von Escherichia coli.
VB198.3.3.2
Das AcrAB-TolC-Effluxpumpensystem von Escherichia coli
6.5.2 Aktin und Myosin im Muskel
6.5.2.1 Aktin und Myosin im Muskel - Konformationsänderungen um Bewegung zu erzeugen
|
|
Bildquelle: 101. Myosinköpfchen |
|
|
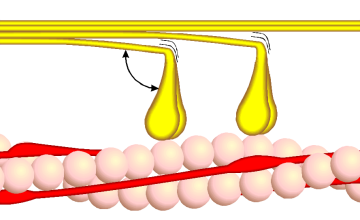
Bildquelle: 80. Querbrückenzyklus Phase 1 - Myosin (gelb) verbindet sich mit Aktin (rosa) unter Aufnahme von Ca++-Ionen. Der angedeutete Winkel beträgt etwa 90°. |

|
|
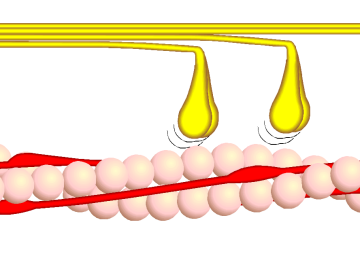
Bildquelle: 81. Querbrückenzyklus Phase 2 - Myosinköpfchen (gelb) kippen und gleiten so am Aktin vorbei (rosa). ATP wird dabei zu ADP und Phosphor verstoffwechselt. Der angedeutete Winkel beträgt etwa 50°. |
|
|
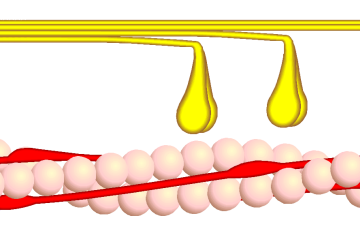
Bildquelle: 82. Querbrückenzyklus Phase 3 - Myosinköpfchen (gelb) lösen sich unter Aufnahme von ATP vom Aktin (rosa). |
|
|
Bildquelle: 83. Querbrückenzyklus Phase 4 - Myosin (gelb) in Ruhezustand. Aktin (rosa). |
|
|
Bildquelle: 99.
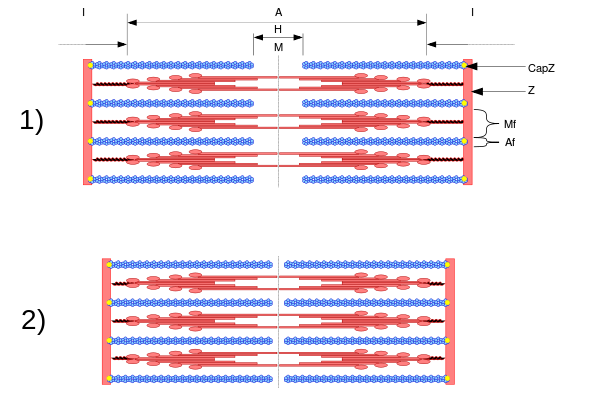
Sarkomer
1) Das Sarkomer, wenn der Muskel gestreckt ist |

|
|
Bildquelle: 97.
Eine elektronenmikroskopische Aufnahme eines quergestreiften Muskels. Der etwas dunklere Streifen ist das A-Band, das oben mit A gekennzeichnet ist. Es hat einen hellen Doppelstrich in der Mitte, der H-Band heißt. Die helleren Streifen heißen I-Band und haben die schwarz erscheinenden Z-Scheiben in der Mitte. Die dunklen Strukturen, die paarweise auf beiden Seiten der Z-Scheiben liegen, sind Mitochondrien, die die Energie für die Arbeit der Muskeln zur Verfügung stellen. |
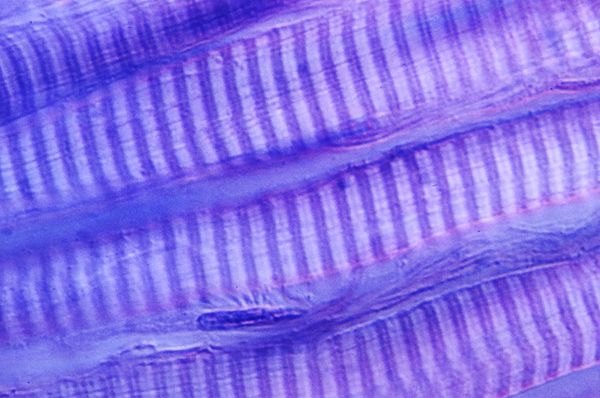
|
|
Bildquelle: 98. Lichtmikroskopischer Längsschnitt quergestreifter Muskelzellen (auch: Muskelfasern) bei starker Vergrößerung (Hämatoxylin-Eosin-Färbung, Interferenzkontrast). |
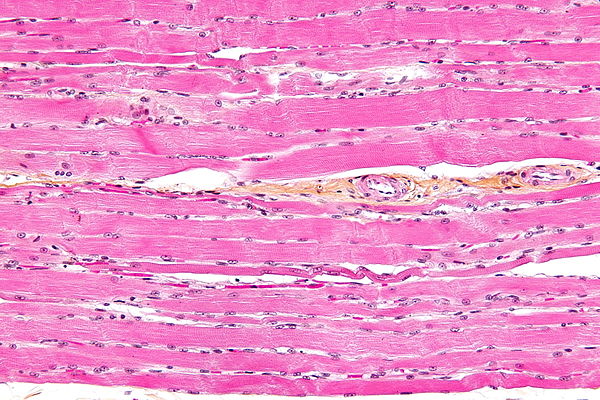
|
|
Bildquelle: 107. Wenn man Muskelfasern in deutlich geringerer Auflösung betrachtet, sieht man daß jede dieser Fasern viele Zellkerne hat, die sich an den Rändern der Zellen befinden. Bei der orangenen Struktur handelt es sich um eine Muskelfascie, die verschiedene Teile des Muskels voneinander abgrenzt, die rosa Ovale darin sind quer geschnittene Adern. |
6.5.2.2 Titin (auch Connectin) im Muskel
Titin, das ich oben bei der schematischen Abbildung des Sarkomers (↑) schon erwähnt hatte, ein elastisches Protein, das das Aktin-Fliament gestreckt hält und an der Z-Scheiben verankert. Es besteht aus zwei Teilen: einem langen Schwanz, der es mit dem Myosin des A-Bandes verbindet und einem elastischen Teil, der aus bis zu 40 aneinandergereihten soganannten immunoglobulin-ähnlichen Domänen besteht. Diese Domäen heißen so, weil sie an bestimmte Strukturen in Antikörpern erinnern, die auch Immunoglobuline heißen.103.
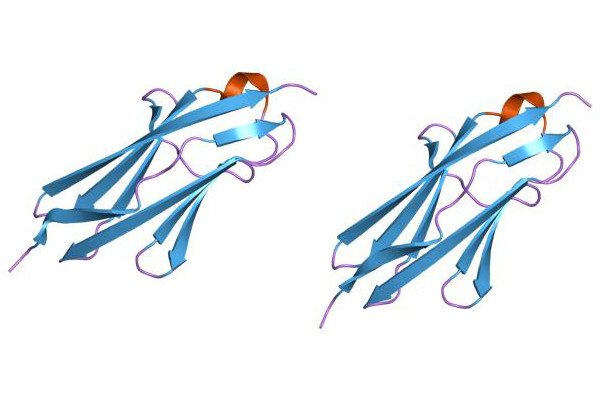
|
|
Bildquelle: 104. Immunoglobulin-ähnliche Domänen von Titin |
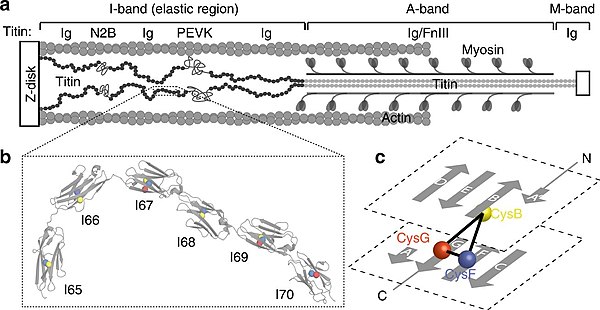
|
|
Bildquelle: 102.1
a - Lage von Titin im Sarkomer |
6.5.3 Aktin und Myosin als wesentlicher Teil des Zellskelettes
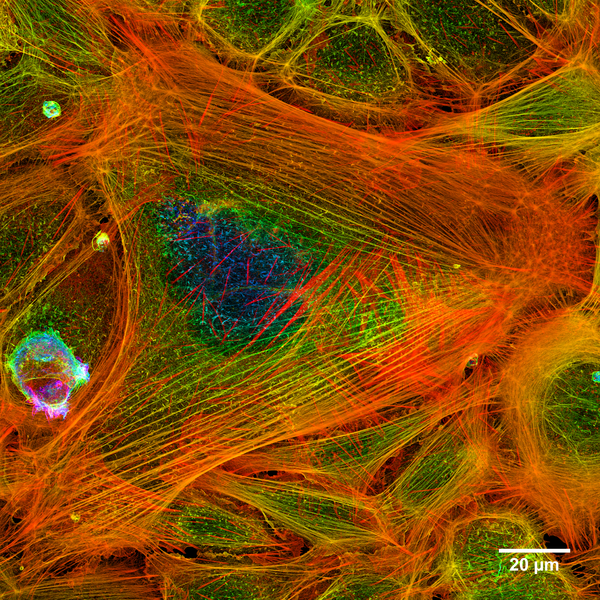
Aktin und Myosin haben im Körper noch diverse andere Funktionen.Aktin bildet einen wesentlichen Teil des Zellskelettes.
|
|
Bildquelle: 85. Aktinfilamente in einer Zelle. Die blau dargestellten Filamente sind am weitesten vorne, dann kommen von vorne nach hinten grün, gelb, orange und rot.
|
6.5.4 Aktin und Myosin in den Mikrovilli
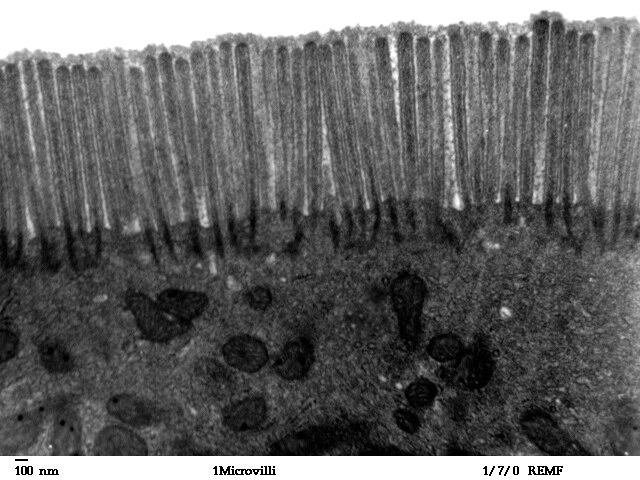
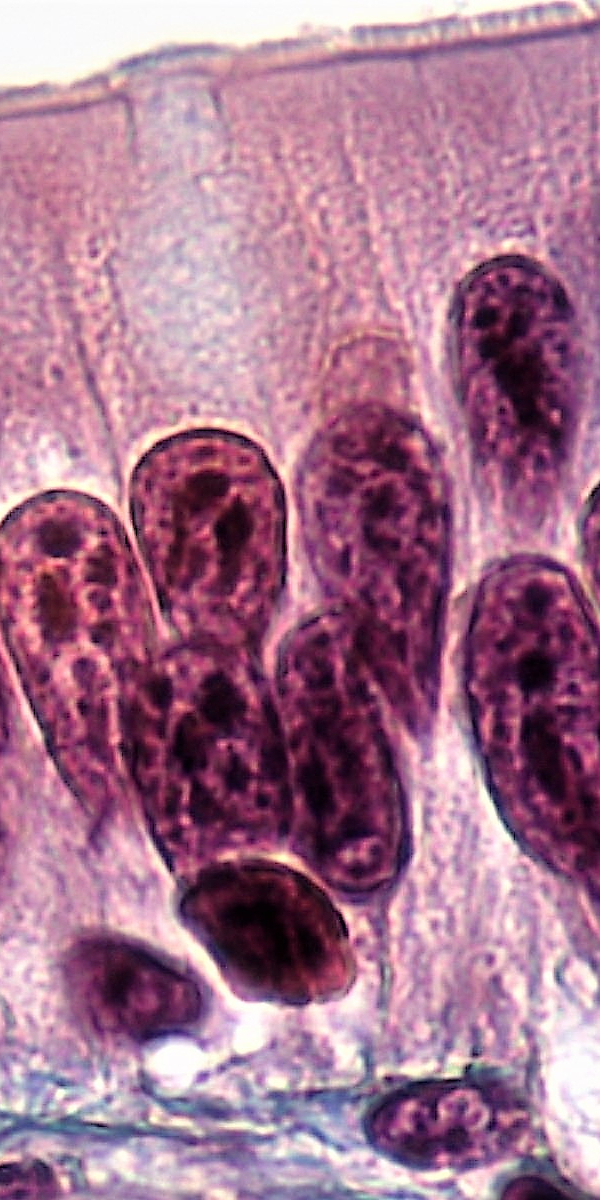
Aktin bildet zum Beispiel in den Mikrovilli ein Filamentbündel, das durch Calmodulin, Myosin, Villin und Fimbrin quervernetzt ist84.. Mikrovilli (Einzahl: Mikrovillus, von lateinisch villus ‚Zotte‘) sind fadenförmige Zellfortsätze, die zur Oberflächenvergrößerung von Zellen und der Verbesserung des Stoffaustausches dienen. Mikrovilli sind hauptsächlich in tierischen Epithelzellen, beispielsweise im Darm, Niere, Geschmacksknospen, in der Gebärmutter und an den Eizellen vorhanden und formen den für diese Epithelien (Oberflächengewebe) typischen Bürstensaum.

|
|
Bildquelle: 84.1 Aktinbündel in den Mikrovilli
|
|
|
Bildquelle: 87. Transmissionselektronenmikroskopsiches Bild von Mikrovilli im menschlichen Dünndarmabschnitt Jejunum |
|
|
Bildquelle: 86. Darmschleimhautzellen, an deren oberen Rand Mikrovilli zu erkennen sind
Weitere Details zum Darm, siehe: |
{kind=link}
6.5.4 Aktin in der Phagozytose
Aktin spielt in der Phagozytose eine wesentliche Rolle. Aktin, Proteine, die Aktin aufbauen, an Aktin binden und Aktinproteine umbauen, bilden den evolutionär konservierten Kern der Phagocytosemaschinerie, den nahezu alle Eukaryoten gemeinsam haben. Die wichtigsten zu diesem Kern gehörenden Proteine heißen actin nucleation complex Arp2/3, seine Regulatoren WASP/N-WASP, WAVE/SCAR, Proteine die aktinfilamente umbauen sind gelsolin, profilin, cofilin, formin und coronin. 109.
6.5.5 Aktin und Scheinfüßchen
lamellipodia and filopodia6.6 Proteine im Gesamtzusammenhang der Zelle: Tubulin und Mikrotubuli
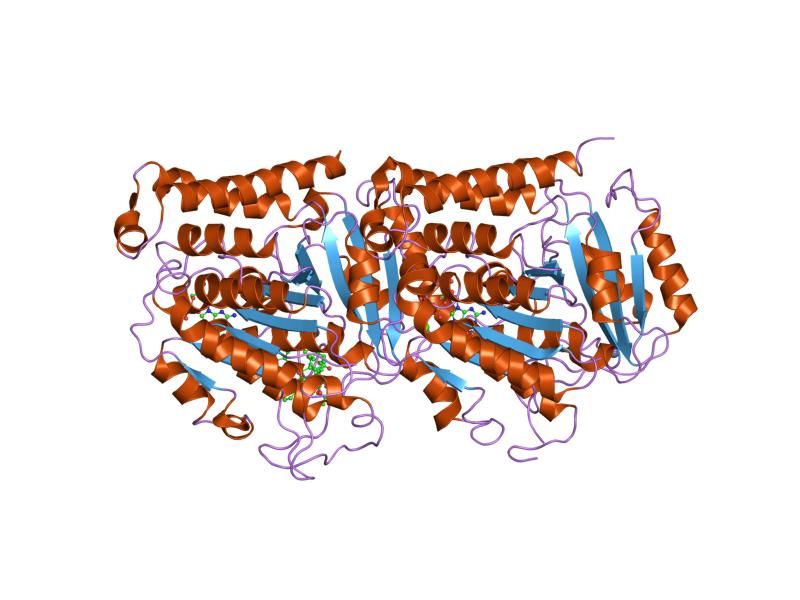
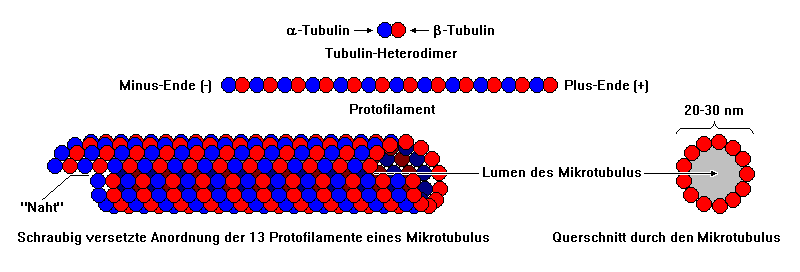
6.6.1 Vom Tubulin zum Mikrotubulus
|
|
Bildquelle: 75. Tubulin-Dimer aus α-Tubulin und β-Tubulin vom Rind (Bos taurus) |
|
|
Bildquelle: 76. Aus α-Tubulin (hier vereinfacht als blaue Kugeln) und β-Tubulin (rote Kugeln) setzen sich Protofilamente zusammen, von denen 13 zusammen einen Mikrotubulus ergeben. |
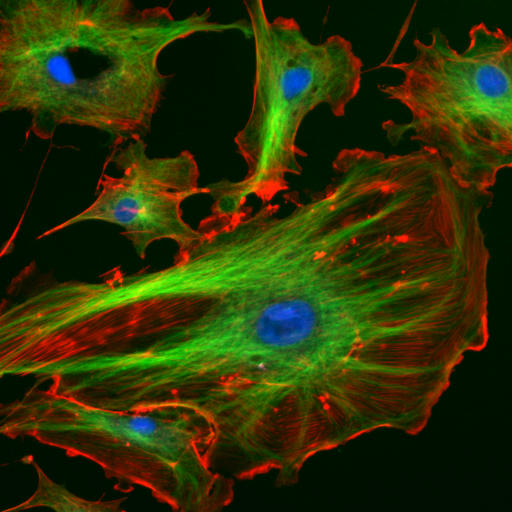
6.6.2 Mikrotubuli als Teil des Zellskelettes
|
|
Bildquelle: 79.
Endothelzellen aus der Inneren Wand (Endothel) von Lungenarterien des Rindes unter dem Mikroskop. Die Zellkerne sind mit DAPI blau markiert. Die Mikrotubuli wurden über einen Antikörper grün markiert. Mit rot fluoreszierendem Phalloidin wurden die Aktinfilamente markiert. |
6.6.3 Mikrotubuli als Eisenbahnschienen der Zelle
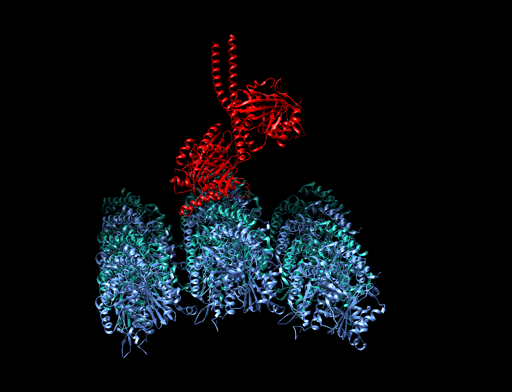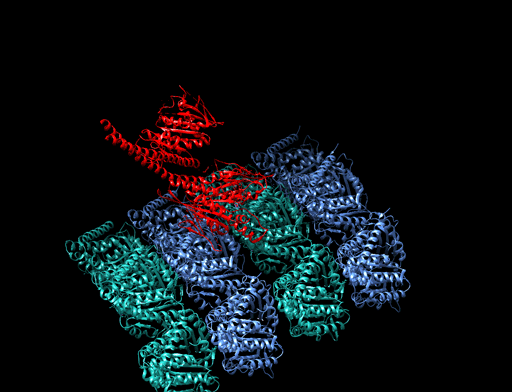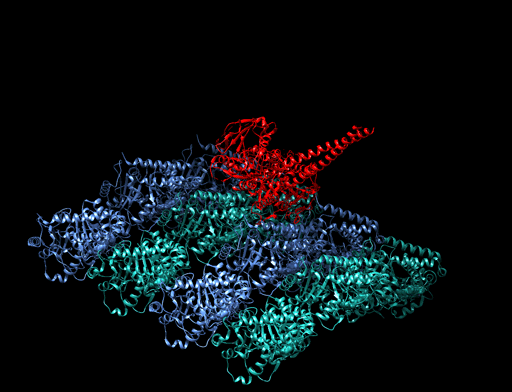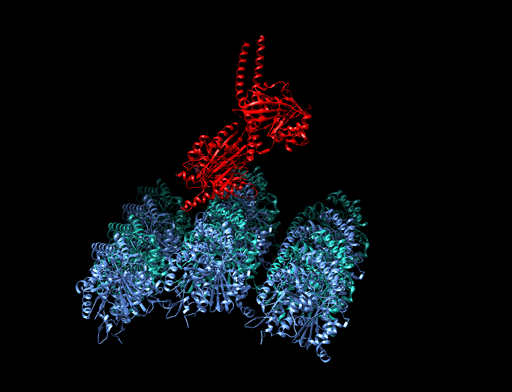
|
|
Bildquelle: 77. Bändermodell von Kinesin auf Tubulin - erkennbar ist, daß die violett und blau dargestellten Tubulinmoleküle durch Kugeln gar nicht so schlecht dargestellt sind und die beiden Füßchen vom Kinesin (rot) tatsächlich ähnlich wie unten dargstellt aussehen. |
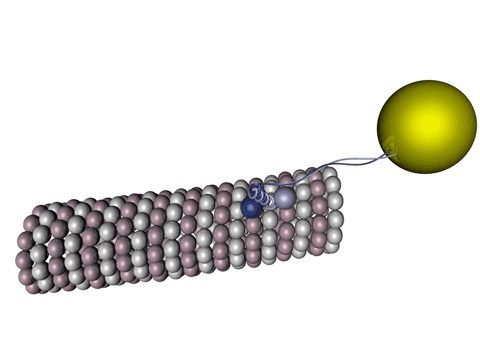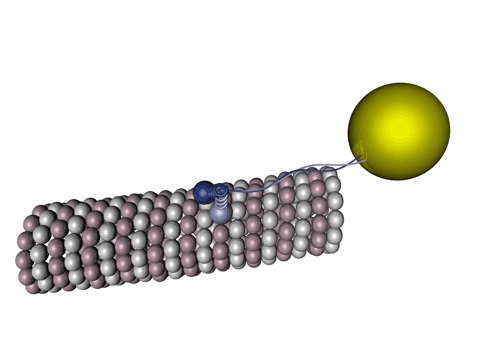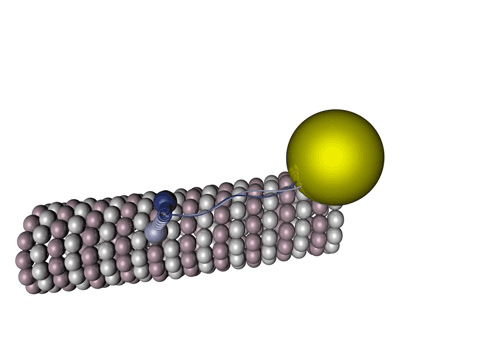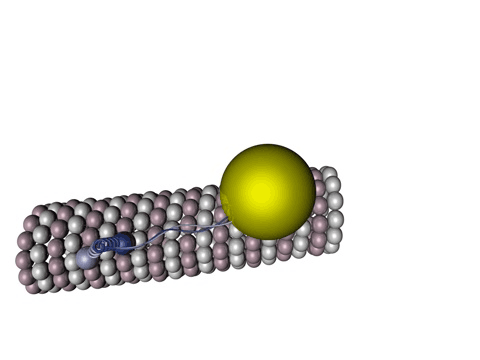
|
|
Bildquelle: 78. Das Motortprotein Kinesin-1 (z. B. KIF5A) bewegt sich entlang eines Mikrotubulus, wobei die einzelnen Köpfe abwechselnd am beta-Tubulin binden. Die gelbe Blase soll das darstellen, was durch den Motor transportiert wird, beispielsweise könnte es ein Bläschen (Vesikel) mit Stoffen sein, die in der Synapse gebraucht werden. |
6.6.4 Mikrotubuli in Zilien und Flagellen
6.6.4.1 Genereller Aufbau von Zilien und Flagellen
Als Flagellen werden zwei völlig verschieden aufgebaute Organellen mit derselben Funktion bezeichnet. Alle Flagellen sind bewegliche haarförmige Gebilde, die die jeweilige Zelle benutzt, um sich fortzubewegen. Die Flagelle der Bakterien ist spiralförmig gedreht ähnlich einem Korkenzieher. Bewegt wird sie mit einem molekularen Motor, der die Flagelle dreht, ähnlich einer Schiffsschraube.112.VB198.3.3.
Das Flagellum der BakterieDie Flagellen der Eukaryoten kennen wir von Spermien und Einzellern. Sie haben eine Struktur aus einem Kreis von zwei mal neun Mikrotubuli, in deren Mitte sich zwei weitere Mikrotubuli befinden. Diese Mikrotubuli bilden die Längsachsen des Skelettes der Flagelle und die Struktur wird als Axonemstruktur bezeichnet (Bauplan 9x2+2).
Als Zilie oder Cilium (lat. cilium ‚Wimper‘, Mehrzahl: cilia) bezeichnet man eine besondere Form des Zellfortsatzes bei Zellen von eukaryotischen Organismen. Es existieren zwei Varianten. Einmal Sekundäre Zilien oder Flimmerhärchen, die wie Flagellen aufgebaut sind und meist beweglich sind (Bauplan 9x2+2). Daneben gibt es auch meist unbewegliche primäre Zilien, in deren Axonemstruktur das mittlere Mikrotubulipaar fehlt (Bauplan 9x2+0). Die primären Zilien haben einerseits mit Sinneswahrnehmungen zu tun, andererseits spielen sie über Zell-zu-Zell-Signale eine wesentliche Rolle in der Embryonalentwicklung.115.
Der Aufbau einer Zilie oder Flagelle beginnt, indem sich ein membranumhüllte Vesikel an die distalen Anghängsel (engl: distal appendages, DAPs) an einem Ende der Mutterzentriole anlagern und dort miteinander verschmelzen, so daß sie dort eine Art Kappe bilden. Die distalen Anhängsel verbinden sich durch die Transitionsfasern mit der Plasmamembran der Zelle und innerhalb dieses so abgegrenzten Bereiches der Membran verschmilzt die Kappe mit der Zellmembran und bilden so den Anfang der Zilienmenbran. Nach und nach werden weitere membranumhüllte Vesikel ergänzt um die so neu entstandene Membran der entstehenden Zilie zu erweitern, so daß sie ausreichend Platz für die wachsende Axonemstruktur bietet. Die so an der Plasmamembran angelagerten Mutterzentriole nennt man den Basalkörper der Zilie. Der Basalkörper besteht aus Mutter- und Tochterzentriolen und enthält auch diverse andere Proteine, von denen viele noch nicht bekannt sind. Zentriolen bestehen aus 9 Gruppen von drei aneinandergelagerten Mikrotubuli (Bauplan 9x3+0). An jeweils zwei dieser drei Mikrotubuli werden innerhalb der Zilienmembran die Axoneme angebaut, die das Rückrat der Mikrotubuli bilden. 116., 118.
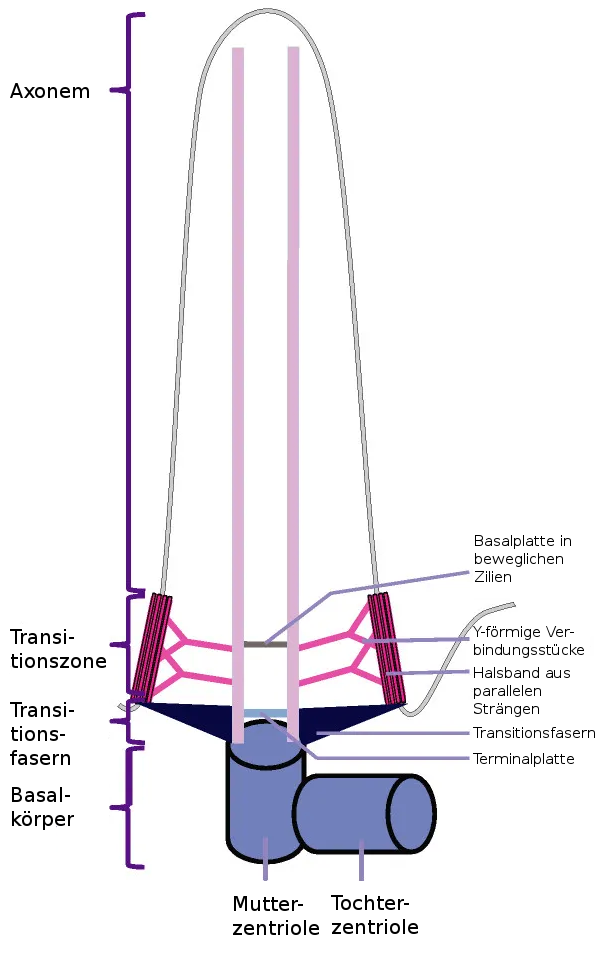
|
|
Bildquelle: 116.1
|
Sehr kleine Proteine können durch die Transitionszone hindurchdiffundieren. Größere können auf unterschiedliche Weise durch die Grenzzone hindurchgeschleust werden. Proteine die an den Motor für den intraflagellaren Transport, das heterotrimere Kinesin II (aus Kif3a, Kif3b, and Kap) gebunden werden, können die Grenzzone passieren. Dies wird durch ciliare Lokalisationssequenzen (englisch: ciliary localization sequences, CLSs) gesteuert, die anzeigen, welche Stoffe für die Zilie bestimmt sind. Mit diesen Signalen werden beispielsweise die für die Zilie bestimmten G-Protein-gekoppelten-Rezeptoren (GPCRs) von ihrem Herstellungsort in die Zilien geschickt. Darüber hinaus können periphere Membranproteine durch spezielle Transportproteine durch die Transitionszone geschleust werden. 117.
Während Kinesin II dem Tranzport zur Spitze des Ziliums dient, wird Zytoplasmisches Dynein 2 benötigt, um Abfallstoffe oder andere für den Zellkörper bestimmte Moleküle aus der Zilie wieder in den Zellkörper zu bevördern.118.
6.6.4.2 Primäre Zilien als Sinnesorgane der Zellen
Bei den primären Zilien fehlen die zentralen inneren Mikrotubuli, sie sind meist nur passiv beweglich und spielen beispielsweise in vielen Sinneszellen eine Rolle110.. Das habe ich am Beispiel des äußeren Segementes des Stäbchens im Auge erklärt.O7.19.2.2.3
Die Funktionsweise des Stäbchens6.6.4.3 Sekundäre Zilien oder Flimmerhärchen
Die Kinozilien oder Flimmerhärchen sind aktiv bewegliche sekundäre Zilien mit zwei zentralen Mikrotubuli und neun Mikrotubulipaaren, die im Kreis darum herum angeordnet sind (Bauplan 9×2+2).
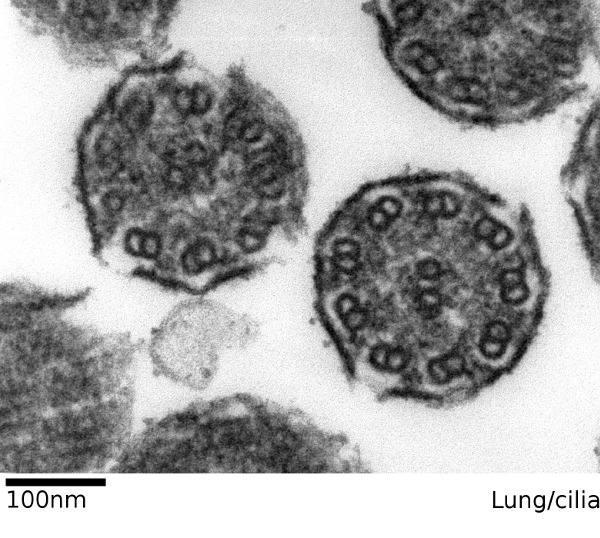
|
|
Bildquelle: 90. Schnitt durch zwei Flimmerhärchen der Lunge eines Säugetiers. Erkennbar sind jeweils zwei Kreisförmige Strukturen in der Mitte und neun Paare an Kreisen außen herum (Bauplan 9×2+2), die Kreise sind Mikrotubuli. |

|
|
Bildquelle: 88. Rasterelektronenmikroskopische Aufnahme des Epithels der Tracheen der Lunge. Dort gibt es Zellen ohne und welche mit Flimmerhärchen. Was wie größere Grasbüschel aussieht, sind Flimmerhärchen. Das dazwischen, was wie ein kurzer Rasen wirkt, sind Mikrovilli. |
|
|
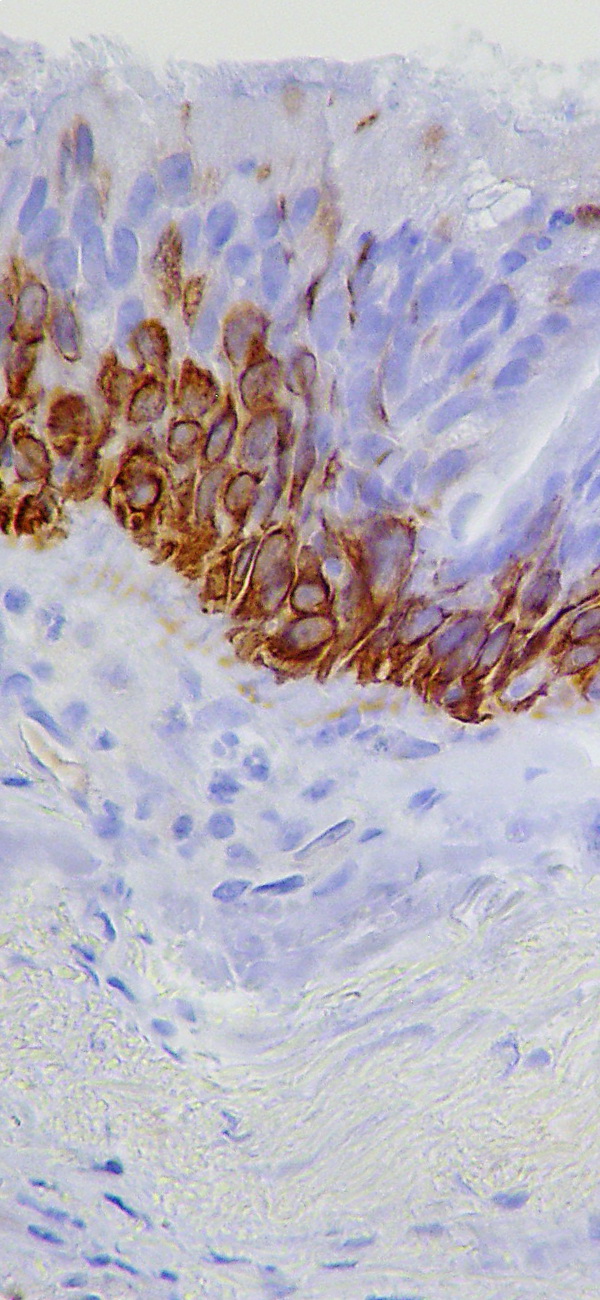
Bildquelle: 89. Normales Epithel der Bronchien (Innenauskleidung der Lungenäste) eines Menschen mit einer Antikörperfärbung gegen Zytokeratin 5 und 6 (CK5-6). Wie das für dieses Epithel normal ist, sind nur die unteren Zellen des Epithels braun angefärbt. Die Flimmerhärchen sind am oberen Rand der obersten Zellen erkennbar. |
6.6.4.3 Die Flagellen der Eukaryoten
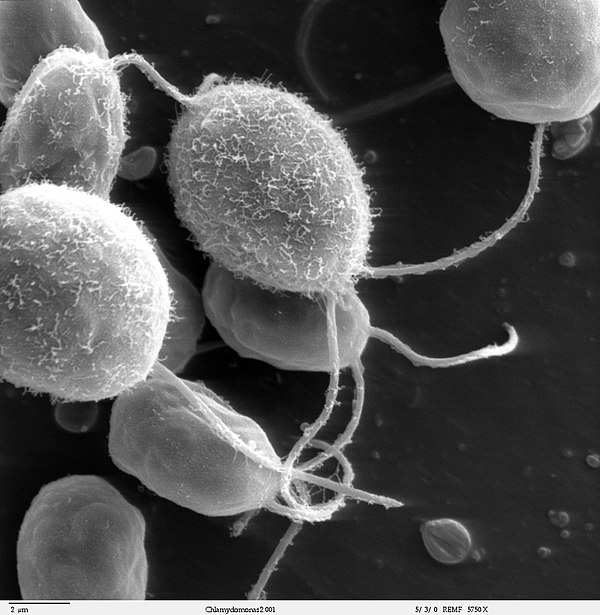
|
|
Bildquelle: 113. Rasterelektronenmikroskopische Aufnahme von Chlamydomanas reinhardtii |
|
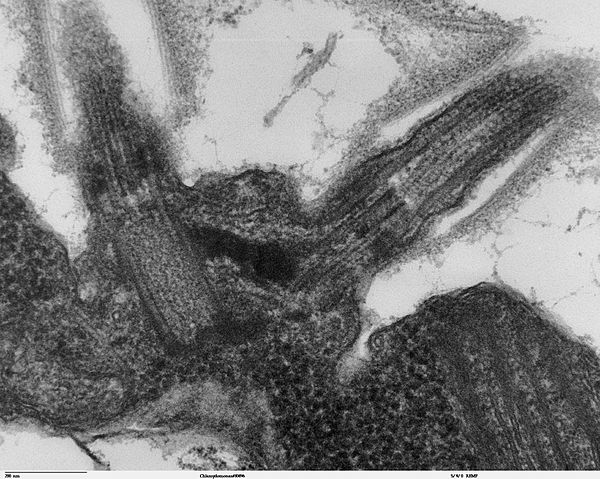
|
Bildquelle: 114. Transmissionselektronenmikroskopisches Bild der Basalkörpers der beiden Flagellen von Chlamydomonas reinhardtii. Diese sind durch ein dunkles Band verbunden von dem angenommen wird, daß es zur Koordanisation der Bewegungen der Flagellen beiträgt. |
6.6.5 Teilungsspindel in der Zellteilung
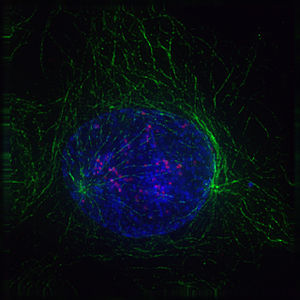
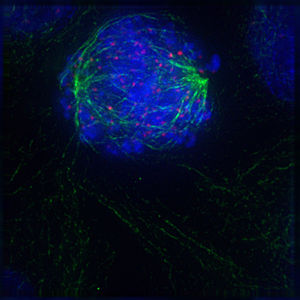
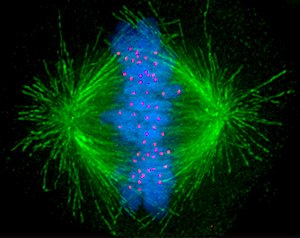
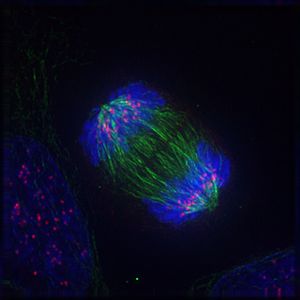
Das Folgende sind fluoreszenzmikroskopische Aufnahmen sich teilender Zellen. Von der Zelle selbst slles was nicht angefärbt wurde, unsichtbar, jedoch ist DNA blau gefärbt, die Kinetochore rot und die Mikrotubuli der Teilungsspindel sind grün gefärbt.
|
|
|
Bildquelle: 91.
Prophase |
Bildquelle: 92.
Prometaphase |
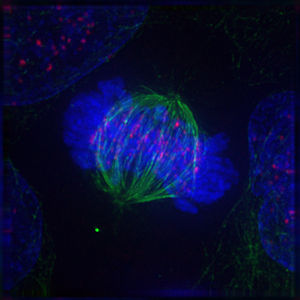
|
|
|
Bildquelle: 93.
Metaphase |
Bildquelle: 94.
Metaphase |
|
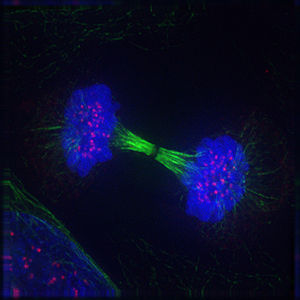
|
|
Bildquelle: 95.
Anaphase |
Bildquelle: 96.
Telophase |
6.6.6
6.6.7 Mikrotubuli und Bewußtsein
VB209.
Quantenprozesse im Gehirn verbinden Körper und Seele7. DNA und RNA
7.1 Nukleoside, die Bausteine von DNA und RNA und der Energiestoffwechsel
7.1.1 Die Grundstrultur der Nukleobasen, Nukleoside und Nucleotide

|
|
Bildquelle: 121.
|
7.1.2 Guanin, Gunanosin und seine Phosphate GDP und GTP
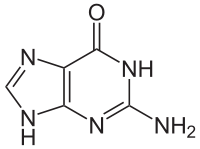
Ein Molekül, das an der Stelle stehen könnte, die im obigen bild der orangene Kasten mit dem Wort Base bezeichnet, ist die Nukleobase Guanin.
|
|
Bildquelle: 122. Guanin |
|
|
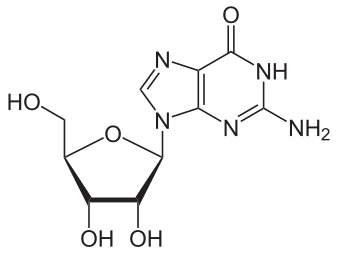
Bildquelle: 123. Guanosin |
|
|
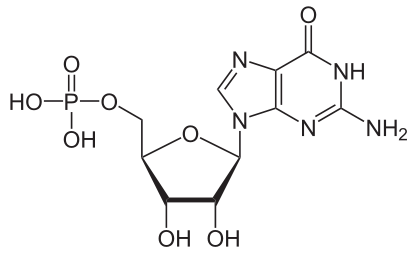
Bildquelle: 124. Guanosinmonophosphat |
|
|
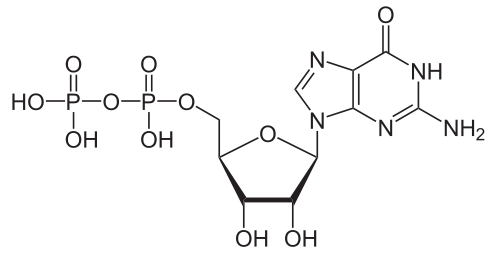
Bildquelle: 125. Guanosindiphosphat (GDP) |
|
|
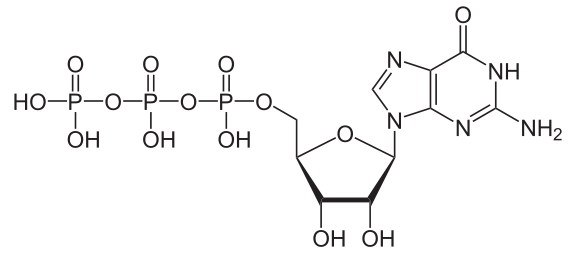
Bildquelle: 126. Guanosintriphosphat (GTP) |
Eine wesentliche Funktion spielen die Nukleotide GDP und GTP auch in der Signalübertragung durch die Zellwand indem die G-Proteine, wenn sie an die G-Protein-Gekoppelten-Rezeptoren gebunden sind, durch GTP in die aktivierte Form überführt werden, die dann andere Proteine aktiviert, wie es beispiesweise das G-Protein Transducin im Auge tut.
O7.19.2.2.3.2.3
Transducin, ein G-Protin
7.1.2 cyclisches Guanosinmonophosphat (cGMP)
|
|
|
Bildquelle: 124. Guanosinmonophosphat (GMP) |
O7.19.2.2.3.6
cGMP und seine Phosphodiesterase (PDE)
|
|
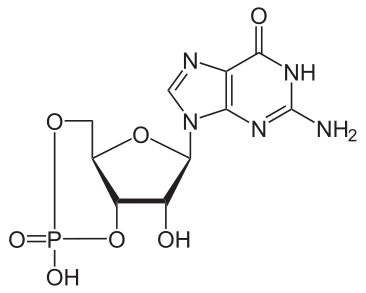
Bildquelle: 129. cyclisches Guanosinmonophosphat (cGMP) |

|
|
Bildquelle: 130. cyclisches Guanosinmonophosphat (cGMP) |
O7.19.2.2.3.2.7
Guanylylcyclasen
7.1.3 Adenin, ADP und ATP
7.1.x Die Basen n der Desoxyribonikleinsäure
|
|
|
Bildquelle: 124. Guanosinmonophosphat |
|
|
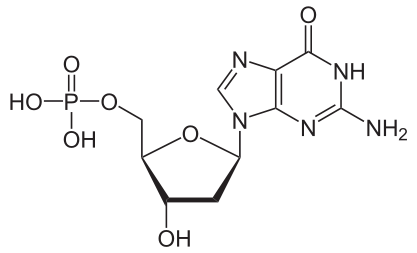
Bildquelle: 127. Desoxyguanosinmonophosphat |
|
|
Bildquelle: 128.
|
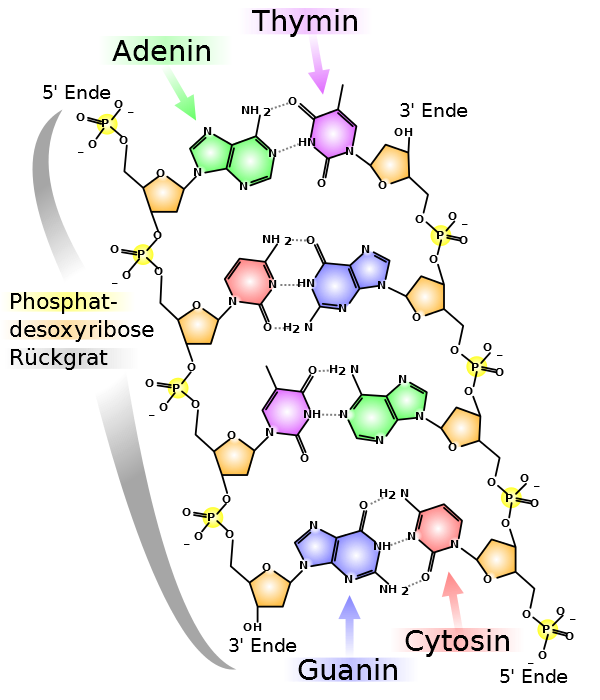
7.2 Darstellungen der DNA
|
|
Bildquelle: 18. Stäbchenmodell der DNA-Doppelhelix |
|
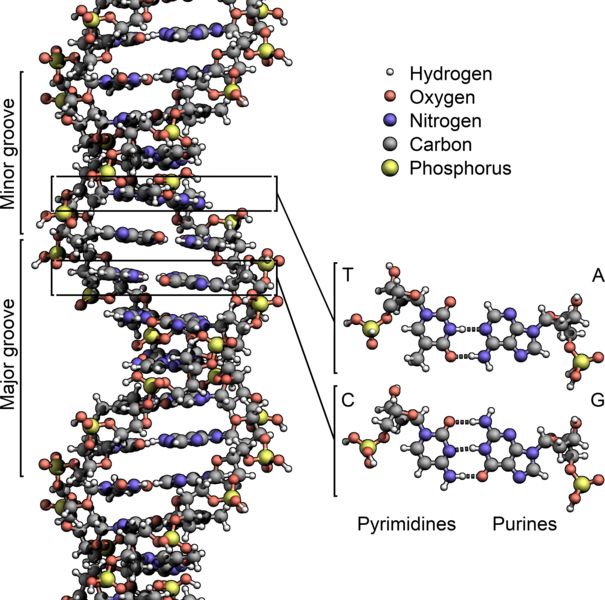
|
Bildquelle: 19.
|
7.3 Verdichtungsgrade der DNA
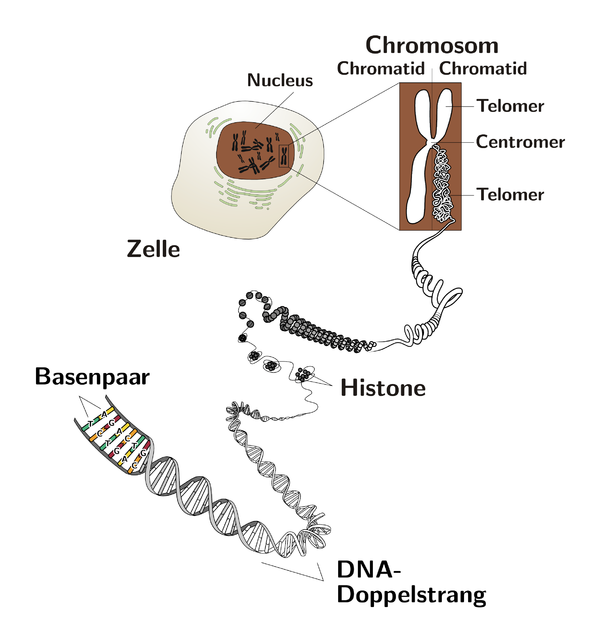
7.3.1 Histone und Nukleosomen
Die DNA liegt aber nicht einfach lose im Zellkern, wie man vielleicht vermuten könnte, sondern sie ist mehr oder weniger aufgewickelt. Der erste Schritt dieses Aufwickelns geschieht mit Proteinen die sich Histone nennen. Die Histone H2A, H2B, H3 und H4 setzten sich zu einem Histon-Oktamer, einem aus acht Teilen bestehenden Proteinkomplex, zusammen, indem von jedem dieser Histone zwei Exemplare eingebaut werden.

|
|
Bildquelle: 21. Bändermodelle der Histone. Rechts oben ist dargestellt, wie die DNA darum herumgewickelt wird, so daß ein Nukleosom entsteht. |
|
|
Bildquelle: 22.
Nukleosom. |
7.3.2 Solenoidstruktur der DNA
Das Histon H1 hat die Funktion dafür zu sorgen, daß diese Perlenkette noch einmal zu einer Spirale aufgerollt wird, zu der Solenoidstruktur. DNA die zu Solenoidstrukturen aufgewickelt ist ist so verdichtet, daß die darauf befindlichen Gene nicht oder selten abgelesen werden. Die Form mit einzelnen Histonen und die Solenoidstruktur sind beides Formen, in denen die DNA normalerweise in der Zelle vorliegt.
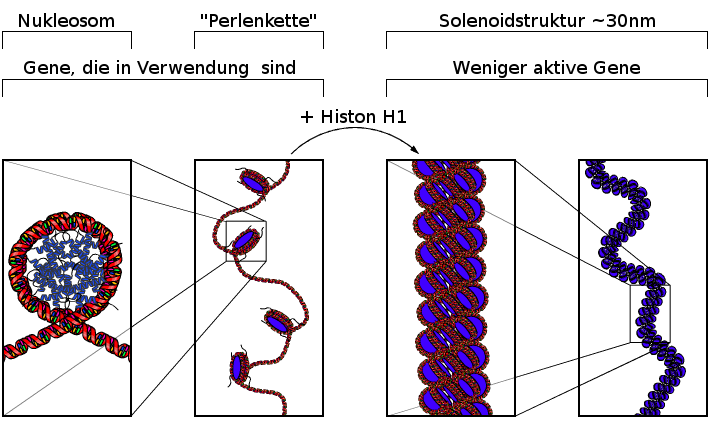
|
|
Bildquelle: 23.
|
7.3.3 Chromosomenterritorien im Zellkern
Die Chromosomen sind aber nicht einfach wild im Zellkern vermischt, sondern jedes Chromosom hat einen eigenen Bereich, sein Chromosomenterritorium.
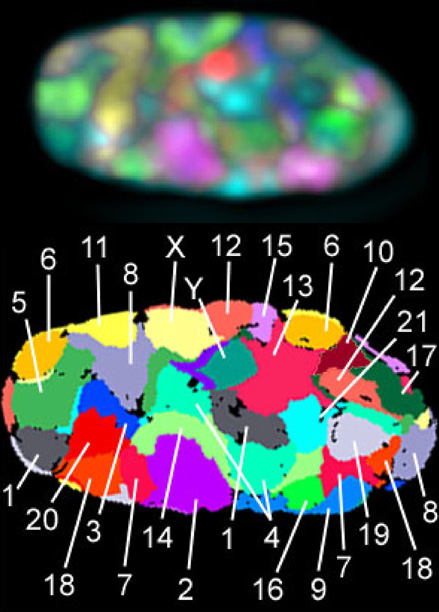
|
|
Bildquelle: 40.
Oben: Zellkern eines menschlichen Fibroblasten, in dem alle 24 verschiedenen Chromosomen (1 - 22, X und Y) per Fluoreszenz in situ Hybridisierung (FISH) mit einer unterschiedlichen Kombination von insgesamt 7 Fluorochromen angefärbt wurden. Gezeigt ist eine mittlere Ebene in einem deconvolvierten Bildstapel, der mit Weitfeld-Mikroskopie aufgenommen wurde. |
|
|
Bildquelle: 41.
|
7.3.4 Chromosomendarstellung in X-Form
Bevor sich eine Zelle teilt, werden die Chromosomen kopiert und die vorher relativ diffus verteilten Chromosomen werden zu unter dem Lichtmikroskop erkennbaren Strukturen zusammengezogen, von denen die meisten wie ein X mit zwei etwas längeren und zwei etwas kürzeren Armen erscheinen.
|
|
Bildquelle: 20.
|
Quellen
-
Bild VB212.PNG:
 File:Holograph-record-de.png auf Wikimedia Commons ist die durch Benutzer:Murkano aus der deutschen Wikipedia erstellte deutsche Version von File:Holography-record.png von User:DrBob aus der Englischen Wikipedia.
File:Holograph-record-de.png auf Wikimedia Commons ist die durch Benutzer:Murkano aus der deutschen Wikipedia erstellte deutsche Version von File:Holography-record.png von User:DrBob aus der Englischen Wikipedia.
Danke das du das Bild unter den Lizenzen GNU 1.2 und CC BY-SA 3.0 freigegeben hast. Thank you very much!
-
siehe auch: Stichwort Holografie in Wikipedia
-
Gerard 't Hooft:
The holographic mapping of the Standard Model onto the black hole horizon, Part I: Abelian vector field, scalar field and BEH Mechanism. In:
 arXiv.org, gr-qc, arXiv:gr-qc/0504120 (Submitted on 25 Apr 2005) ( Volltext)
arXiv.org, gr-qc, arXiv:gr-qc/0504120 (Submitted on 25 Apr 2005) ( Volltext)
-
↑Bild VB218.PNG:
File:Benzene Representations-numbers.svg von User:Vladsinger (Original) und User:Armando-Martin (Nummerierung) beide von Wikimedia Commons
Vielen Dank daß Du das Bild unter GNU 1.2, CC BY-SA 3.0 hochgeladen hast! Thank you very much!
-
↑Bild VB218.JPG: Ausschnitt aus
File:Fale - Monaco - 86.jpg von Fabio Alessandro Locati (=User:Fale von Wikimedia Commons), beschnitten durch Kersti Nebelsiek
Vielen Dank daß Du das Bild unter CC BY-SA 3.0 hochgeladen hast! Thank you very much!
-
↑Bild VB21801.JPG:
File:Cristal de roca Soria.jpg von User:Bergminer von Wikimedia Commons
Vielen Dank daß Du das Bild unter CC BY-SA 4.0 hochgeladen hast! Thank you very much!
-
↑Bild VB21801.PNG:
File:Periodic table (German).svg von User:Dr.cueppers von Wikimedia Commons
Vielen Dank daß Du das Bild völlig freigegeben hast! Thank you very much!
-
↑Bild VB21802.JPG:
File:Double slit x-ray simulation trans-long 07500 eV.jpg von User:Timm Weitkamp von Wikimedia Commons
Vielen Dank daß Du das Bild unter CC BY-SA 3.0 hochgeladen hast! Thank you very much!
-
↑Bild VB21803.JPG:
File:PSM V13 D178 Superposition of two wave systems.jpg aus: Norman Lockyer: Water waves and sound waves. In: The Popular science monthly, Vol. VIII, Issue 9, June 1878, S.166-173
-
↑Bild VB21804.JPG:
File:Swimming Pool Interferometry.jpg und hier von M. Alexander, European Southern Observatory (ESO)
Vielen Dank, daß Sie das Bild unter CC BY-SA 4.0 hochgeladen baben! Thank you very much!
-
↑Bild VB21802.PNG:
File:53 iodine (I) enhanced Bohr model.png von User:Ahazard.sciencewriter von Wikimedia Commons
Vielen Dank daß Du das Bild unter CC BY-SA 4.0 hochgeladen hast! Thank you very much!
-
↑Bild VB218.GIF:
File:Bohr atom animation 2.gif von User:Kurzon von Wikimedia Commons
Vielen Dank daß Du das Bild unter CC BY-SA 3.0 hochgeladen hast! Thank you very much!
-
↑Bild VB21801.GIF:
Von Kersti Nebelsiek aus den folgenden Bildern zusammengesetzt: File:Espectro H absorción.GIF von User:Juancarcole und File:Emission spectrum-H labeled.svg von User:Adrignola, User:Merikanto von Wikimedia Commons
Lizenz: CC BY-SA 3.0, Danke! Thank you very much!
-
↑Bild VB21803.PNG:
File:Atomic-orbital-clouds spd m0.png von User:Geek3 von Wikimedia Commons
Vielen Dank daß Du das Bild unter CC BY-SA 4.0 hochgeladen hast! Thank you very much!
-
↑Bild VB201.PNG:
File:Flageolette.svg von User:Mjchael, Danke, daß Du das Bild unter der Lizenz CC BY-SA 2.5 zur Verfügung gestellt hast. Thank you very much!
-
↑Bild VB20101.JPG:
File:Chladni pattern 4.jpg von User:Elmar Bergeler von Wikimedia Commons
Vielen Dank daß Du das Bild unter CC BY-SA 3.0 hochgeladen hast! Thank you very much!
-
↑Bild VB21804.PNG:
600 Pixel PNG-Version von: File:EisenatomLichteffekt.svg das User:Groogokk, aufbauend auf Bildern von User:Fornax und User:Halfdan (alle von Wikimedia Commons) erstellt hat.
Vielen Dank daß Du das Bild unter CC BY-SA 3.0 hochgeladen hast! Thank you very much!
-
↑Bild V0272.PNG:
File:DNA Structure+Key+Labelled.pn NoBB.png von User:Zephyris (Richard Wheeler) von Wikimedia Commons
Vielen Dank daß Du das Bild unter GNU 1.2, CC BY-SA 3.0 hochgeladen hast! Thank you very much!
-
↑Bild VB21805.PNG:
File:Difference DNA RNA-DE.svg von User:Sponk von Wikimedia Commons
Vielen Dank daß Du das Bild unter GNU 1.2, CC BY-SA 3.0 hochgeladen hast! Thank you very much!
-
↑Bild VB21806.PNG:
File:Chromosom und DNA.png vom National Human Genome Research Institute, deutsche Übersetzung: User:San Jose von Wikimedia Commons
Dieses Bild steht unter public domain da es Inhalte enthält, die die National Institutes of Health der USa erstellt haben.
-
↑Bild VB21807.PNG:
File:Nucleosome structure-2.png von Richard Wheeler (User:Zephyris), changes User:Rekymanto, beide von Wikimedia Commons
Vielen Dank daß Du das Bild unter GNU 1.2, CC BY-SA 3.0 hochgeladen hast! Thank you very much!
-
↑Bild VB21802.GIF:
File:Nucleosome core particle 1EQZ large.gif von User:Darekk2 von Wikimedia Commons, nach den Daten von Proteinstuktuer 1EQZ, RCSB PDB aus Joel M. Harp, Leif Hanson, David E. Timmc, Gerard J. Bunicka: Asymmetries in the nucleosome core particle at 2.5 A resolution. In: Acta crystallographica. Section D, Biological crystallography, 2000 Dec;56(Pt 12):1513-34 PMID: 11092917 ( Volltext)
Vielen Dank daß Sie das Bild unter CC BY-SA 3.0 hochgeladen haben! Thank you very much!
-
↑Bild VB21808.PNG: Ausschnitt aus
Bild V027303.PNG: File:Chromatin Structures.png von Richard Wheeler (User:Zephyris) von Wikimedia Commons
Vielen Dank daß Du das Bild unter GNU 1.2, CC BY-SA 3.0 hochgeladen hast! Thank you very much!
-
James A. Perkins:
A History of Molecular Representation. Part One: 1800 to the 1960s. In: Journal of Biocommunication (JBC) ( Volltext)
-
Robert B. Corey,
Linus Pauling: Molecular Models of Amino Acids, Peptides, and Proteins. In: Review of Scientific Instruments 24, 621 (1953) ( Volltext)
-
Brittanica-Autoren,
William L. Hosch, Amy Tikkanen, Aakanksha Gaur: Stichwort: Electron diffraction ( Volltext) in BVI.6 Encyclopaedia Britannica., abgerufen 1.11.2019
-
Walter L. Koltun:
Precision space-filling atomic models. In: Biopolymers, Volume3, Issue6, December 1965, Pages 665-679 https://doi.org/10.1002/bip.360030606
-
Matteo Leone,
Nadia Robotti: Frédéric Joliot, Irène Curie and the early history of the positron (1932–33). In: European Journal of Physics 31 (2010) 975–987 doi:10.1088/0143-0807/31/4/027 ( Volltext)
-
Dieter Meschede:
B143.2
Gerthsen Physik. (21. Auflage 2002) Berlin, Heidelberg: Springer Verlag ISBN 3-540-42024-X
- 29.1 Auf den vorderen Umschlagseiten. Entgegen den üblichen Geflogenheiten habe ich die Zahlen so umgerechnet, daß sie alle mit dem Faktor *10-31kg multipliziert werden, weil dann einfacher auf den ersten Blick zu sehen ist, wie groß der Unterschied zwischen den Werten ist.
-
Tilo Fischer,
Hans-Jerg Dorn:
B143.3
Physikalische Formeln und Daten. (1982) Stuttgart: Klett Schulbuchverlag GmbH, ISBN 3-12-770800-9
-
Georg Pfotzer:
Antiproton und Antineutron. In: Physikalische Blätter, Volume 13, Issue 4, April 1957, Pages 152–164 ( Volltext)
-
Bild VA309.PNG:
File:Paarbildung gamma p Desy Blasenkammer Rekonstruiert left.png
von
Ivan Baev (Benutzer:Schroedinger),
GNU-Lizenz für freie Dokumentation, Version 1.2 und CC BY-SA 3.0
-
Frank R. N. Gurd:
The use of Corey-Pauling-Koltun space-filling models in teaching. In: Biochemical Education, Volume2, Issue2, April 1974, Pages 27-29 ( Volltext)
-
Clinton Joseph Davisson,
Lester Halbert Germer: Diffraction of Electrons by a Crystal of Nickel. In: Physical Review, Vol. 30, Iss. 6 — December 1927 ( Volltext)
-
George Paget Thomson,
Alexander Reid: Diffraction of Cathode Rays by a Thin Film.In: Nature 119, 890 (1927) doi:10.1038/119890a0
-
Alexander Reid:
The diffraction of cathode rays by thin celluloid films. In: Proceedings of the Royal Society of London A, 2 July 1928, Volume 119, Issue 783 ( Volltext)
-
Henning Sievers:
Louis de Broglie und die Quantenmechanik. In: arXiv.org (1998) arXiv:physics/9807012 ( Volltext)
-
Stepan S. Batsanov:
Van der Waals Radii of Elements. BZ300. Inorganic Materials, Vol. 37, No. 9, 2001, pp. 871–885. Translated from Neorganicheskie Materialy, Vol. 37, No. 9, 2001, pp. 1031–1046. ( Volltext)
-
William Lawrence Bragg:
The structure of some crystals as indicated by their diffraction of X-rays. In: Proceedings of the Royal Society of London, A, 22 September 1913, Volume 89, Issue 610 ( Volltext 1, 2)
-
Bild VB21805.JPG:
File:PLoSBiol3.5.Fig1bNucleus46Chromosomes.jpg, Teil von Figure 1 aus Andreas Bolzer, Gregor Kreth, Irina Solovei, Daniela Koehler, Kaan Saracoglu, Christine Fauth, Stefan Müller, Roland Eils, Christoph Cremer, Michael R. Speicher, Thomas Cremer: Three-Dimensional Maps of All Chromosomes in Human Male Fibroblast Nuclei and Prometaphase Rosettes. In: PLOS Biology, April 26, 2005, 3(5): e157. ( Volltext)
Vielen Dank, daß Sie das Bild unter CC BY 2.5 hochgeladen haben! Thank you very much!
-
Bild VB21806.JPG:
File:Nuclear Architecture.pdf von Evin Wieser (User:Ewieser94) von Wikimedia Commons
Vielen Dank, daß Du das Bild unter CC BY-SA 4.0 hochgeladen hast! Thank you very much!
-
Somi Kim,
Nam-Kyung Yu, Bong-Kiun Kaang: CTCF as a multifunctional protein in genome regulation and gene expression. In: Experimental & Molecular Medicine, volume 47, page e166(2015) ( Volltext)
-
Jesse R. Dixon,
Siddarth Selvaraj, Feng Yue, Audrey Kim, Yan Li, Yin Shen, Ming Hu, Jun S. Liu, Bing Ren: Topological Domains in Mammalian Genomes Identified by Analysis of Chromatin Interactions. In: Nature. 2012 May 17; 485(7398): 376–380. ( Volltext)
-
↑Bild VB21809.PNG:
File:Aminosaeuren.png von Benutzer:MarkusZi aus der deutschen Wikipedia
Vielen Dank, daß Du das Bild unter GNU 1.2, CC BY-SA 3.0 hochgeladen hast! Thank you very much!
-
↑Bild VB21810.PNG:
File:AminoAcidball.svg von User:YassineMrabet von Wikimedia Commons
Vielen Dank, daß Du das Bild unter vollständig freigegeben hast! Thank you very much!
-
↑Bild VB21811.PNG:
File:Tetrapeptide structural formulae v.1.png von User:Jü von Wikimedia Commons
Vielen Dank, daß Du das Bild unter vollständig freigegeben hast! Thank you very much!
-
Bild VB21807.JPG:
File:StrukturaSekondare.jpg von User:A.Jashari von Wikimedia Commons
Vielen Dank, daß Du das Bild unter CC BY-SA 3.0 hochgeladen hast! Thank you very much!
-
↑Bild VB21812.PNG:
File:Protein IFNA2 PDB 1itf.png von User:Emw von Wikimedia Commons
Vielen Dank, daß Du das Bild unter GNU 1.2, CC BY-SA 3.0 hochgeladen hast! Thank you very much!
-
↑Bild VB21813.PNG:
File:Beta sheet bonding antiparallel-color.svg von User:Fvasconcellos von Wikimedia Commons
Vielen Dank, daß Du das Bild als gemeinfrei veröffentlich hast! Thank you very much!
-
↑Bild VB21814.PNG:
File:Beta sheet bonding parallel-color.svg von User:Fvasconcellos von Wikimedia Commons
Vielen Dank, daß Du das Bild als gemeinfrei veröffentlich hast! Thank you very much!
-
↑Bild VB21815.PNG:
File:Beta-Faltblatt.svg von User:Roland.chem von Wikimedia Commons
Vielen Dank, daß Du das Bild unter CC0 1.0 hochgeladen hast! Thank you very much!
-
Bild VB21808.JPG:
File:Antiparallell-beta-pleated-sheet.jpg von Användare:Vili der schwedischen Wikipedia
Vielen Dank, daß Du das Bild unter GNU 1.2, CC BY-SA 3.0 hochgeladen hast! Thank you very much!
-
Bild VB21809.JPG:
File:Parallell-beta-pleated-sheet.jpg von Användare:Vili der schwedischen Wikipedia
Vielen Dank, daß Du das Bild unter GNU 1.2, CC BY-SA 3.0 hochgeladen hast! Thank you very much!
-
↑Bild VB21816.PNG:
File:Feuillets bêta.svg von Utilisateur:Pou24 ais der französischen Wikipedia
Vielen Dank, daß Du das Bild unter CC BY 1.0 hochgeladen hast! Thank you very much!
-
↑Bild VB21817.PNG:
File:GFP structure.png von Richard Wheeler (User:Zephyris) von Wikimedia Commons
Vielen Dank, daß Du das Bild unter GNU 1.2, CC BY-SA 3.0 hochgeladen hast! Thank you very much!
-
Mats Ormö,
Andrew B. Cubitt, Karen Kallio, Larry A. Gross, Roger Y. Tsien, S. James Remington: Crystal Structure of the Aequorea victoria Green Fluorescent Protein. In: Science, 06 Sep 1996: Vol. 273, Issue 5280, pp. 1392-1395, DOI: 10.1126/science.273.5280.1392 PMID: 8703075 ( Volltext)
-
↑Bild VB21818.PNG:
File:Beta schleife.svg von User:Muskid von Wikimedia Commons
Vielen Dank, daß Du das Bild wegen zu geringer Schöpfungshöhe als gemeinfrei hochgeladen hast! Thank you very much!
-
↑Bild VB21819.PNG:
File:1m8n Choristoneura fumiferana.png von User:WillowW aus der englischen Wikipedia
Vielen Dank, daß Du das Bild unter GNU 1.2, CC BY-SA 3.0 hochgeladen hast! Thank you very much!
-
↑Bild VB21820.PNG:
File:1ezg Tenebrio molitor.png von User:WillowW von Wikimedia Commons
Vielen Dank, daß Du das Bild unter GNU 1.2, CC BY-SA 3.0 hochgeladen hast! Thank you very much!
-
Bild VB21810.JPG:
File:PDB 1ncj EBI.jpg und hier von Jawahar Swaminathan und MSD Mitarbeiter am Europäischen Institut für Bioinformatik
Vielen Dank, daß Sie das Bild als gemeinfrei veröffentlicht haben! Thank you very much!
-
Gregory M. Martin,
Craig Yoshioka, Emily A. Rex, Jonathan F. Fay, Qing Xie, Matthew R. Whorton, James Z. Chen, Show-Ling Shyng: Cryo-EM structure of the ATP-sensitive potassium channel illuminates mechanisms of assembly and gating. In: eLife 2017;6:e24149 doi: 10.7554/eLife.24149 ( Volltext)
- 61.1 Bild VB21811.JPG:
File:Elife-24149-fig2-v2.jpg aus Gregory M. Martin, Craig Yoshioka, Emily A. Rex, Jonathan F. Fay, Qing Xie, Matthew R. Whorton, James Z. Chen, Show-Ling Shyng: Cryo-EM structure of the ATP-sensitive potassium channel illuminates mechanisms of assembly and gating. In: eLife 2017;6:e24149 doi: 10.7554/eLife.24149 ( Volltext)
Vielen Dank, daß Sie das Bild unter CC BY-SA 4.0 hochgeladen haben! Thank you very much!
- 61.2 Bild VB21812.JPG:
File:Elife-24149-fig3-v2.jpg aus Gregory M. Martin, Craig Yoshioka, Emily A. Rex, Jonathan F. Fay, Qing Xie, Matthew R. Whorton, James Z. Chen, Show-Ling Shyng: Cryo-EM structure of the ATP-sensitive potassium channel illuminates mechanisms of assembly and gating. In: eLife 2017;6:e24149 doi: 10.7554/eLife.24149 ( Volltext)
Vielen Dank, daß Sie das Bild unter CC BY-SA 4.0 hochgeladen haben! Thank you very much!
- 61.3
Bild VB21813.JPG:
File:Elife-24149-fig7-v2.jpg aus Gregory M. Martin, Craig Yoshioka, Emily A. Rex, Jonathan F. Fay, Qing Xie, Matthew R. Whorton, James Z. Chen, Show-Ling Shyng: Cryo-EM structure of the ATP-sensitive potassium channel illuminates mechanisms of assembly and gating. In: eLife 2017;6:e24149 doi: 10.7554/eLife.24149 ( Volltext)
Vielen Dank, daß Sie das Bild unter CC BY-SA 4.0 hochgeladen haben! Thank you very much!
- 61.4
Bild VB21814.JPG:
File:Elife-24149-fig8-v2.jpg aus Gregory M. Martin, Craig Yoshioka, Emily A. Rex, Jonathan F. Fay, Qing Xie, Matthew R. Whorton, James Z. Chen, Show-Ling Shyng: Cryo-EM structure of the ATP-sensitive potassium channel illuminates mechanisms of assembly and gating. In: eLife 2017;6:e24149 doi: 10.7554/eLife.24149 ( Volltext)
Vielen Dank, daß Sie das Bild unter CC BY-SA 4.0 hochgeladen haben! Thank you very much!
- 61.5
Bild VB21822.PNG: Abwandlung eines Ausschnittes aus
Bild VB21814.JPG, File:Elife-24149-fig8-v2.jpg aus Gregory M. Martin, Craig Yoshioka, Emily A. Rex, Jonathan F. Fay, Qing Xie, Matthew R. Whorton, James Z. Chen, Show-Ling Shyng: Cryo-EM structure of the ATP-sensitive potassium channel illuminates mechanisms of assembly and gating. In: eLife 2017;6:e24149 doi: 10.7554/eLife.24149 ( Volltext)
Vielen Dank, daß Sie das Bild unter CC BY-SA 4.0 hochgeladen haben! Thank you very much!
- 61.6
Bild VB21821.PNG: Ausschnitt aus
Bild VB21811.JPG: File:Elife-24149-fig2-v2.jpg aus Gregory M. Martin, Craig Yoshioka, Emily A. Rex, Jonathan F. Fay, Qing Xie, Matthew R. Whorton, James Z. Chen, Show-Ling Shyng: Cryo-EM structure of the ATP-sensitive potassium channel illuminates mechanisms of assembly and gating. In: eLife 2017;6:e24149 doi: 10.7554/eLife.24149 ( Volltext)
Vielen Dank, daß Sie das Bild unter CC BY-SA 4.0 hochgeladen haben! Thank you very much!
- 61.1 Bild VB21811.JPG:
-
Bild VB21804.GIF:
File:CrawlingCelegans.gif oder hier von Bob Goldstein
Vielen Dank, daß Sie das Bild unter GNU 1.2, CC BY-SA 3.0 hochgeladen haben! Thank you very much!
-
Marcus Winkler:
B114.8
Struktur-Funktions-Untersuchungen an ATP-empfindlichen K+-Kanälen, deren Untereinheiten und schließenden Liganden. (2009) Dissertation der Fakultät für Chemie und Pharmazie der Eberhard Karls Universität Tübingen zur Erlangung des Grades eines Doktors der Naturwissenschaften ( Volltext)
-
Bild VB21823.PNG:
File:Glucose Insulin Release Pancreas.svg von User:Aydintay von Wikimedia Commons
Vielen Dank, daß Du das Bild als gemeinfrei hochgeladen hast! Thank you very much!
-
Masaru Emoto
(aus dem Japanischen von Dr. Monika Wacker):
B124.1.1
Wasserkristalle. Was das Wasser zu sagen hat. (2003) Burgrain: Koha Verlag. ISBN 3-929512-20-3
-
Masaru Emoto
(aus dem Japanischen von Dr. Monika Wacker):
B124.1.2
Die Antwort des Wassers. (2002) Burgrain: Koha Verlag. ISBN 3-929512-93-9
-
Masaru Emoto
(aus dem Englischen von Urs Thoenen):
B124.1.3
Die Botschaft des Wassers. Sensationelle Bilder von gefrorenen Wasserkristallen. Band 1.(2002) Burgrain: Koha Verlag. ISBN 3-929512-21-1
-
Bild VB21815.JPG:
File:H2O-Tetraeder.jpg von Benutzer:Solid State der Deutschen Wikipedia
Vielen Dank, daß Du das Bild unter GNU 1.2, CC BY-SA 3.0 hochgeladen hast! Thank you very much!
-
Bild VB21824.PNG:
File:Watermolecule.svg von Patrick-Emil Zörner (User:Paddy) und User:Sgbeer von Wikimedia Commons
Vielen Dank, daß ihr das Bild unter CC BY-SA 2.0 hochgeladen habt! Thank you very much!
-
Bild VB21825.PNG:
File:3D model hydrogen bonds in water.svg von Wikipedista:Qwerter aus der Tchechischen Wikipedia, mit Anpassungen von User:Snek01 von Wikimedia Commons
Vielen Dank, daß Du das Bild unter CC BY-SA 3.0 hochgeladen hast! Thank you very much!
-
Bild VB21826.PNG:
File:冰晶结构.png von User:IgniX von Wikimedia Commons
Vielen Dank, daß Du das Bild unter GNU 1.2, CC BY-SA 3.0 hochgeladen hast! Thank you very much!
-
Bild VB21827.PNG:
File:冰晶结构2.png von User:IgniX von Wikimedia Commons
Vielen Dank, daß Du das Bild unter GNU 1.2, CC BY-SA 3.0 hochgeladen hast! Thank you very much!
-
Bild VB21828.PNG:
Schneeflockenfotos von Wilhelm Bentley, die von Kersti Nebelsiek zu einem Gesamtbild zusammengesetzt wurden
Die Einzelbilder sind aufgrund ihres Alters Gemeinfrei, daher gebe ich das Gesamtbild ebenfalls vollständig frei
-
Bild VB21816.JPG:
File:Snowflake macro photography 1 (cropped).jpg von User:Alexey Kljatov von Wikimedia Commons
Vielen Dank, daß Du das Bild unter CC BY-SA 4.0 hochgeladen hast! Thank you very much!
-
Bild VB21817.JPG:
File:PDB 1jff EBI.jpg von Jawahar Swaminathan und MSD Mitarbeiter am Europäischen Institut für Bioinformatik
Vielen Dank, daß ihr das Bild unter Public Domain hochgeladen habt! Thank you very much!
-
Bild VB21829.PNG:
File:Mikrotubula007 de.png von User:Qniemiec von Wikimedia Commons
Vielen Dank, daß Du das Bild unter GNU 1.2, CC BY-SA 4.0, CC BY-SA 3.0, CC BY-SA 2.5, CC BY-SA 2.0, CC BY-SA 1.0 hochgeladen hast! Thank you very much!
-
Bild VB20901.GIF:
File:KinMT Gif.gif von User:Kaden.Rawson von Wikimedia Commons
Vielen Dank daß Du das Bild unter CC BY-SA 4.0 hochgeladen hast! Thank you very much!
-
Bild VB209.GIF:
File:Mechanismus der Bewegung eines Kinesinmoleküls entlang eines Mikrotubulus .gif von Konrad J. Böhm (Benutzer:Kboehm auf Wikipedia)
Danke, daß Du das Bild unter CC BY-SA 4.0 hochgeladen hast! Thank you very much!
-
Bild VB21819.JPG:
File:FluorescentCells.jpg
Dieses Werk ist in den Vereinigten Staaten gemeinfrei, da es von Mitarbeitern der US-amerikanischen Bundesregierung oder einem ihrer Organe in Ausübung ihrer dienstlichen Pflichten erstellt wurde
-
↑Bild VB21830.PNG:
File:Querbrückenzyklus 1.png von Benutzer:Moralapostel aus der deutschen Wikipedia
Vielen Dank, daß Du das Bild unter GNU 1.2, CC BY-SA 3.0 hochgeladen hast! Thank you very much!
-
↑Bild VB21831.PNG:
File:Querbrückenzyklus 2.png von Benutzer:Moralapostel aus der deutschen Wikipedia
Vielen Dank, daß Du das Bild unter GNU 1.2, CC BY-SA 3.0 hochgeladen hast! Thank you very much!
-
↑Bild VB21832.PNG:
File:Querbrückenzyklus 3.png von Benutzer:Moralapostel aus der deutschen Wikipedia
Vielen Dank, daß Du das Bild unter GNU 1.2, CC BY-SA 3.0 hochgeladen hast! Thank you very much!
-
↑Bild VB21833.PNG:
File:Querbrückenzyklus 4.png von Benutzer:Moralapostel aus der deutschen Wikipedia
Vielen Dank, daß Du das Bild unter GNU 1.2, CC BY-SA 3.0 hochgeladen hast! Thank you very much!
-
Jeffrey W. Brown,
C. James McKnight: Molecular Model of the Microvillar Cytoskeleton and Organization of the Brush Border. In: PLoS One February 24, 2010, doi.org/10.1371/journal.pone.0009406 ( Volltext)
- 84.1
↑Bild VB21834.PNG:
File:Microvillar citoskeleton.png aus Jeffrey W. Brown,
C. James McKnight: Molecular Model of the Microvillar Cytoskeleton and Organization of the Brush Border. In: PLoS One February 24, 2010, doi.org/10.1371/journal.pone.0009406 ( Volltext)
Vielen Dank, daß Du das Bild unter CC BY-SA 3.0 hochgeladen hast! Thank you very much!
- 84.1
↑Bild VB21834.PNG:
-
↑Bild VB21835.PNG:
File:STD Depth Coded Stack Phallodin Stained Actin Filaments.png von User:Methylated603 von Wikimedia Commons
Vielen Dank, daß Du das Bild unter CC BY-SA 4.0 hochgeladen hast! Thank you very much!
-
↑Bild VB21820.JPG:
Ausschnitt aus File:Epithelial Tissues Brush Border in Simple Columnar Epithelium (27854453998).jpg und hier, aus der Berkshire Community College Bioscience Image Library
Diese Datei wird unter der Creative-Commons-Lizenz „CC0 1.0 Verzicht auf das Copyright“ zur Verfügung gestellt. Vielen Dank! Thank you very much!
-
↑Bild VB21821.JPG:
File:Microvilli.jpg und hier von Louisa Howard, Katherine Connolly - Dartmouth Electron Microscope Facility
Public domain Dieses Werk wurde von seinem Urheber Dartmouth Electron Microscope Facility als gemeinfrei veröffentlicht. Dies gilt weltweit. Vielen Dank! Thank you very much!
-
↑Bild VB21822.JPG:
File:Bronchiolar epithelium 3 - SEM.jpg von seinem Urheber Charles Daghlian als gemeinfrei veröffentlicht
Vielen Dank! Thank you very much!
-
↑Bild VB21823.JPG:
File:Normal bronchial epithelium - CK5-6 (5762353448).jpg oder hier von Yale Rosen (Flickr User)
Vielen Dank, daß Sie das Bild unter CC BY-SA 2.0 hochgeladen haben! Thank you very much!
-
↑Bild VB21824.JPG:
File:Bronchiolar area cilia cross-sections 2.jpg von Louisa Howard, Michael Binder
Dieses Werk wurde von seinem Urhebern als gemeinfrei veröffentlicht. Dies gilt weltweit. Vielen Dank! Thank you very much!
-
↑Bild VB21825.JPG:
File:ProphaseIF.jpg von Roy van Heesbeen (User:Royvanheesbeen) von Wikimedia Commons
Dieses Werk wurde von seinem Urheber als gemeinfrei veröffentlicht. Dies gilt weltweit. Vielen Dank! Thank you very much!
-
↑Bild VB21826.JPG:
File:Prometaphase.jpg von Roy van Heesbeen (User:Royvanheesbeen) von Wikimedia Commons
Dieses Werk wurde von seinem Urheber als gemeinfrei veröffentlicht. Dies gilt weltweit. Vielen Dank! Thank you very much!
-
↑Bild VB21827.JPG:
File:MetaphaseIF.jpg von Roy van Heesbeen (User:Royvanheesbeen) von Wikimedia Commons
Dieses Werk wurde von seinem Urheber als gemeinfrei veröffentlicht. Dies gilt weltweit. Vielen Dank! Thank you very much!
-
↑Bild VB21828.JPG:
File:Kinetochore.jpg von User:Afunguy von Wikimedia Commons
Dieses Werk wurde von seinem Urheber als gemeinfrei veröffentlicht. Dies gilt weltweit. Vielen Dank! Thank you very much!
-
↑Bild VB21829.JPG:
File:Anaphase IF.jpg von Roy van Heesbeen (User:Royvanheesbeen) von Wikimedia Commons
Dieses Werk wurde von seinem Urheber als gemeinfrei veröffentlicht. Dies gilt weltweit. Vielen Dank! Thank you very much!
-
↑Bild VB21827.JPG:
File:TelophaseIF.jpg von Roy van Heesbeen (User:Royvanheesbeen) von Wikimedia Commons
Dieses Werk wurde von seinem Urheber als gemeinfrei veröffentlicht. Dies gilt weltweit. Vielen Dank! Thank you very much!
-
↑Bild VB21831.JPG:
File:Electronic microscopy picture of a muscle.jpg von User:Giolicc von Wikimedia Commons
Vielen Dank, daß Du das Bild unter CC BY-SA 4.0 hochgeladen hast! Thank you very much!
-
↑Bild VB21832.JPG:
File:Muskel quergestreift.JPG von User:Rollroboter von Wikimedia Commons
Vielen Dank, daß Du das Bild unter CC BY-SA 3.0 hochgeladen hast! Thank you very much!
-
↑Bild VB21837.PNG:
File:Sarcomere diagram2.svg, Original von David Richfield (User:Slashme), Beschriftung: User:Marek Mazurkiewicz, beide von Wikimedia Commons
Vielen Dank, daß Du das Bild unter GNU 1.2, CC BY-SA 3.0 hochgeladen hast! Thank you very much!
-
Edward C. Eckels,
Shubhasis Haldar, Rafael Tapia-Rojo, Jaime Andrés Rivas-Pardo, Julio M. Fernández: The Mechanical Power of Titin Folding. In: Cell Reports, Volume 27, Issue 6, 7 May 2019, Pages 1836-1847.e4 ( Volltext 1, 2)
-
↑Bild VB21838.PNG:
File:018-Myosin-1b7t.tiff aus: David Goodsell: Myosin. Molecular motors fueled by ATP power the contraction of muscles ( Volltext)
Vielen Dank, daß Sie das Bild unter CC BY-SA 4.0 hochgeladen haben! Thank you very much!
-
David Giganti,
Kevin Yan, Carmen L. Badilla, Julio M. Fernández, Jorge Alegre-Cebollada: Disulfide isomerization reactions in titin immunoglobulin domains enable a mode of protein elasticity. In: Nature Communications, volume 9, Article number: 185 (2018) ( Volltext)
- 102.1
↑Bild VB21833.JPG:
File:Titin IG Domains.jpg aus: David Giganti, Kevin Yan, Carmen L. Badilla, Julio M. Fernández, Jorge Alegre-Cebollada: Disulfide isomerization reactions in titin immunoglobulin domains enable a mode of protein elasticity. In: Nature Communications, volume 9, Article number: 185 (2018) ( Volltext)
Vielen Dank, daß Du das Bild unter CC BY-SA 4.0 hochgeladen hast! Thank you very much!
- 102.1
↑Bild VB21833.JPG:
-
Olga Mayans,
Jochen Wuerges, Santiago Canela, Mathias Gautel, Matthias Wilmanns: Structural Evidence for a Possible Role of Reversible Disulphide Bridge Formation in the Elasticity of the Muscle Protein Titin. In: Structure, Volume 9, ISSUE 4, P331-340, April 01, 2001 ( Volltext)
-
↑Bild VB21834.JPG:
File:PDB 1g1c EBI.jpg von Jawahar Swaminathan und MSD Mitarbeiter am Europäischen Institut für Bioinformatik, Proteinstruktur nach PDBe › 1g1c
Diese Bild- oder Mediendatei wurde von ihrem Urheber zur uneingeschränkten Nutzung freigegeben. Das Bild ist damit gemeinfrei („public domain“). Dies gilt weltweit. Vielen Dank! Thank you very much!
-
Cristina Machadoa,
Deborah J. Andrew: D-Titin. A Giant Protein with Dual Roles in Chromosomes and Muscles. In: Journal of Cell Biology (JCB), 2000 Oct 30; 151(3): 639–652. ( doi: 10.1083/jcb.151.3.639, Volltext)
-
Cristina Machadoa,
Claudio E. Sunkel, Deborah J. Andrew: Human Autoantibodies Reveal Titin as a Chromosomal Protein. In: Journal of Cell Biology (JCB), 20. April 1998, 141 (2): 321, DOI: 10.1083/jcb.141.2.321 ( Volltext)
-
↑Bild VB21835.JPG:
File:Skeletal striated muscle.jpg von User:Nephron von Wikimedia Commons
Vielen Dank, daß Du das Bild unter GNU 1.2, CC BY-SA 3.0 hochgeladen hast! Thank you very much!
-
Kiisa C. Nishikawa,
Jenna A. Monroy, Theodore E. Uyeno, Sang Hoon Yeo, Dinesh K. Pai, Stan L. Lindstedt: Is titin a ‘winding filament’? A new twist on muscle contraction. In: Proceedings of the Royal Society of London, B 2012 Mar 7; 279(1730): 981–990. ( Volltext)
-
Natalya Yutin,
Maxim Y. Wolf, Yuri I. Wolf, Eugene V. Koonin: The origins of phagocytosis and eukaryogenesis. BZ329. Biology direct, 2009; 4: 9. Published online 2009 Feb 26. doi: 10.1186/1745-6150-4-9 ( Volltext)
-
Hemant Khanna:
Photoreceptor Sensory Cilium: Traversing the Ciliary Gate. In: Cells 2015, 4(4), 674-686; ( Volltext)
-
Hyuk Wan Ko:
The primary cilium as a multiple cellular signaling scaffold in development and disease. In: BMB Reports, Volume 45 Issue 8 / Pages.427-432 / 2012 / 1976-670X(eISSN) ( Volltext)
-
Thierry Mora,
Howard Yu, Yoshiyuki Sowa, Ned S. Wingreen: Steps in the Bacterial Flagellar Motor. In: PLOS Computational Biology, 5(10) 2009 ( Volltext)
-
↑Bild VB21836.JPG:
File:Chlamydomonas2-1.jpg von Wikimedia Commons
Dieses Werk wurde von seinem Urheber Dartmouth Electron Microscope Facility, Dartmouth College als gemeinfrei veröffentlicht. Vielen Dank! Thank you very much!
-
↑Bild VB21837.JPG:
File:Chlamydomonas TEM 08.jpg von Wikimedia Commons
Dieses Werk wurde von seinem Urheber Dartmouth Electron Microscope Facility, Dartmouth College als gemeinfrei veröffentlicht. Vielen Dank! Thank you very much!
-
Nathalie Falk,
Marlene Lösl, Nadja Schröder, Andreas Gießl: Specialized Cilia in Mammalian Sensory Systems. In: Cells 2015, 4(3), 500-519; ( Volltext)
-
Katarzyna Szymanska,
Colin A. Johnson: The transition zone: an essential functional compartment of cilia. In: Cilia, volume 1, Article number: 10 (2012) ( Volltext)
- 116.1 ↑Bild VB21839.PNG:
Übersetzter Ausschnitt aus Figure 1 aus Katarzyna Szymanska,
Colin A. Johnson: The transition zone: an essential functional compartment of cilia. In: Cilia, volume 1, Article number: 10 (2012) ( Volltext)
Vielen Dank, daß Sie das Bild unter CC BY 2.0 hochgeladen haben! Thank you very much!
- 116.1 ↑Bild VB21839.PNG:
Übersetzter Ausschnitt aus
-
Francesc R. Garcia-Gonzalo,
Jeremy F. Reiter: Open Sesame: How Transition Fibers and the Transition Zone Control Ciliary Composition. In: Cold Spring Harbor perspectives in biology 2017 Feb; 9(2): a028134. doi: 10.1101/cshperspect.a028134 ( Volltext)
-
Huihui Yang,
Kaiyao Huang: Dissecting the Vesicular Trafficking Function of IFT Subunits. In: Frontiers in Cell and Developmental Biology, January 2020, Volume 7, Article 352 ( Volltext)
-
↑Bild VB21840.PNG:
File:GPCR structure and receptor.svg von User:Fred the Oyster von Wikimedia Commons
Vielen Dank, daß Du das Bild unter GNU 1.2, CC BY-SA 4.0, CC BY-SA 3.0, CC BY-SA 2.5, CC BY-SA 2.0, CC BY-SA 1.0 hochgeladen hast! Thank you very much!
-
Peter Werner Hildebrand,
Robert Preissner, Cornelius Frömmel: Structural features of transmembrane helices. In: FEBS Letters, Volume 559, Issues 1–3, 13 February 2004, Pages 145-151 ( Volltext)
-
↑Bild VB21841.PNG:
File:Nucleotide nucleoside general.svg von User:Yikrazuul von Wikimedia Commons
Vielen Dank, daß Du das Bild unter CC BY-SA 3.0 hochgeladen hast! Thank you very much!
-
↑Bild VB21842.PNG:
File:Guanin.svg von User:NEUROtiker von Wikimedia Commons
Diese Datei ist gemeinfrei („public domain“), weil sie nur Allgemeingut enthält und die nötige Schöpfungshöhe nicht erreicht.
-
↑Bild VB21842.PNG:
File:Guanosin.svg von User:NEUROtiker von Wikimedia Commons
Diese Datei ist gemeinfrei („public domain“), weil sie nur Allgemeingut enthält und die nötige Schöpfungshöhe nicht erreicht.
-
↑Bild VB21844.PNG:
File:Guanosinmonophosphat protoniert.svg von User:NEUROtiker von Wikimedia Commons
Diese Datei ist gemeinfrei („public domain“), weil sie nur Allgemeingut enthält und die nötige Schöpfungshöhe nicht erreicht.
-
↑Bild VB21845.PNG:
File:Guanosindiphosphat protoniert.svg von User:NEUROtiker von Wikimedia Commons
Diese Datei ist gemeinfrei („public domain“), weil sie nur Allgemeingut enthält und die nötige Schöpfungshöhe nicht erreicht.
-
↑Bild VB21846.PNG:
File:Guanosintriphosphat protoniert.svg von User:NEUROtiker von Wikimedia Commons
Diese Datei ist gemeinfrei („public domain“), weil sie nur Allgemeingut enthält und die nötige Schöpfungshöhe nicht erreicht.
-
↑Bild VB21847.PNG:
File:Desoxyguanosinmonophosphat protoniert.svg von User:NEUROtiker von Wikimedia Commons
Diese Datei ist gemeinfrei („public domain“), weil sie nur Allgemeingut enthält und die nötige Schöpfungshöhe nicht erreicht.
-
↑Bild VB21848.PNG":
File:Chemische Struktur der DNA.svg von Madeleine Price Ball (User:Madprime) Text: Matt (User:Matthias M.), beide von Wikimedia Commons
Vielen Dank, daß Du das Bild unter GNU 1.2, CC BY-SA 3.0, CC BY-SA 2.5, CC BY-SA 2.0, CC BY-SA 1.0 hochgeladen hast! Thank you very much!
-
↑Bild VB21849.PNG:
File:CGMP.svg von User:NEUROtiker von Wikimedia Commons
Diese Datei ist gemeinfrei („public domain“), weil sie nur Allgemeingut enthält und die nötige Schöpfungshöhe nicht erreicht.
-
↑Bild VB21850.PNG:
File:CGMP.png von David Iberri (User:Diberri aus der englischen Wikipedia)
Vielen Dank, daß Du das Bild unter GNU 1.2, CC BY-SA 3.0 hochgeladen hast! Thank you very much!
-
A. Keith Dunker,
David J. Zaleske: Stereochemical considerations for constructing alpha-helical protein bundles with particular application to membrane proteins. In: The Biochemical journal, 1977 Apr 1; 163(1): 45–57. doi: 10.1042/bj1630045, PMID: 869919 ( Volltext)
-
↑Bild O00071910.PNG:
Abwandung von Figure 1. aus Joseph T. Ortega,
Beata Jastrzebska: The Retinoid and Non-Retinoid Ligands of the Rod Visual G Protein-Coupled Receptor. In: International Journal of Molecular Sciences 2019, 20, 6218; doi:10.3390/ijms20246218 ( Volltext 1, 2), umgedreht, neu beschriftet und Membran eingezeichnet durch Kersti Nebelsiek
Vielen Dank, daß Sie das Bild unter CC BY 4.0 hochgeladen haben! Thank you very much!
-
Doug Marman:
The Lenses of Perception Interpretation of Quantum Mechanics. In: Integral Review, August 2018, Vol. 14, No. 1 ( Volltext)
-
↑Bild VB22620.PNG:
File:LSD-3D-vdW.png von User:Benjah-bmm27 von Wikimedia Commons
Vielen Dank, daß Du das Bild als gemeinfrei hochgeladen hast! Thank you very much!
-
↑Bild VB21838.JPG:
Ausschnitt aus File:Choristoneura fumiferana.jpg ( oder hier) vom USDA Forest Service - Northeastern Area Archive, USDA Forest Service, United States
CC BY 3.0 Vielen Dank! Thank you very much!
-
↑Bild VB21839.JPG:
File:Choristoneura fumiferana eggs.jpg ( oder hier) vom USDA Forest Service - Northeastern Area , USDA Forest Service, Bugwood.org
CC BY 3.0 Vielen Dank! Thank you very much!
-
↑Bild VB21840.JPG:
File:Choristoneura fumiferana larva.jpg ( oder hier) von Jerald E. Dewey, USDA Forest Service, Bugwood.org
CC BY 3.0 Vielen Dank! Thank you very much!
-
Michael J. Kuiper,
Craig J. Morton, Sneha E. Abraham, Angus Gray-Weale: The biological function of an insect antifreeze protein simulated by molecular dynamics. In: eLife 4:e05142. May 7, 2015 ( Volltext)
-
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
.svg){kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
_enhanced_Bohr_model.png){kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
.jpg){kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
.jpg){kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
.jpg){kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}